Studenții, studenții absolvenți, tinerii oameni de știință care folosesc baza de cunoștințe în studiile și munca lor vă vor fi foarte recunoscători.
postat pe http://www.allbest.ru/
postat pe http://www.allbest.ru/
MINISTERUL EDUCAȚIEI AL REPUBLICII BELARUS
instituție educațională
„STATUL MOZYR
UNIVERSITATEA PEDAGOGICA ei. I.P. SHAMYAKIN"
DEPARTAMENTUL DE BIOLOGIE
DEPARTAMENTUL GESTIONAREA NATURII SI PROTECTIA NATURII
Cursuri pe disciplină
„fiziologia plantelor”
Influența mineralelor asupra creșterii și dezvoltării plantelor
Executor testamentar:
Bogdanovich Vladimir Grigorievici
MOZYR 2011
INTRODUCERE
CAPITOLUL 1. REVISTA LITERATURĂ
1.3 Fosfor
1,6 calciu
1,7 magneziu
3.4 Deficit de azot
3.5 Deficit de fosfor
3.6 Deficit de sulf
3.7 Deficit de potasiu
3.8 Deficit de calciu
3.9 Deficit de magneziu
CONCLUZIE
REFERINȚE
INTRODUCERE
minerale vegetale
Nutriția minerală a plantelor - un set de procese de absorbție, mișcare și asimilare de către plante a elementelor chimice obținute din sol sub formă de ioni de săruri minerale.
Fiecare element chimic joacă un rol special în viața unei plante.
Azotul este un component al aminoacizilor, elementele de bază care formează proteinele. Azotul este inclus și în mulți alți compuși: purine, alcaloizi, enzime, regulatori de creștere, clorofilă și membrane celulare.
Fosforul este absorbit de plantă sub formă de săruri ale acidului fosforic (fosfați) și se găsește în el în stare liberă sau împreună cu proteinele și alte substanțe organice care alcătuiesc plasma și nucleul.
Sulful este absorbit de plantă sub formă de săruri de acid sulfuric, face parte din proteine și uleiuri esențiale.
Potasiul este concentrat în organele tinere bogate în plasmă, precum și în organele de acumulare a substanțelor de rezervă - semințe, tuberculi, joacă probabil rolul de neutralizator al reacției acide a sevei celulare și este implicat în turgor.
Magneziul se găsește în plantă în același loc cu potasiul și, în plus, face parte din clorofilă.
Calciul se acumulează în organele adulte, în special în frunze, servește ca neutralizator al acidului oxalic dăunător plantei și o protejează de efectele toxice ale diferitelor săruri și participă la formarea membranelor mecanice.
Pe lângă aceste elemente vitale, de o anumită importanță sunt clorura de sodiu, manganul, fierul, fluorul, iodul, bromul, zincul, cobaltul, care stimulează creșterea plantelor.
Scop: Studierea efectului mineralelor asupra creșterii și dezvoltării plantelor.
1. Să studieze materialul despre principalele tipuri de minerale și efectul acestora asupra creșterii și dezvoltării plantelor.
2. Familiarizați-vă cu metodele de determinare a substanțelor minerale din țesuturile plantelor.
3. Identificați simptomele de conținut mineral insuficient și excesiv la plante
CAPITOLUL 1. REVISTA LITERATURĂ
Plantele sunt capabile să absoarbă din mediu în cantități mai mari sau mai mici aproape toate elementele tabelului periodic. Între timp, pentru ciclul normal de viață al unui organism vegetal, este necesar doar un anumit grup de elemente nutritive de bază, ale căror funcții în plantă nu pot fi înlocuite cu alte elemente chimice. Acest grup include următoarele 19 elemente:
Dintre acești nutrienți principali, doar 16 sunt de fapt minerali, deoarece C, H și O pătrund în plante în principal sub formă de CO 2 , O 2 și H 2 O. Elementele Na, Si și Co sunt date între paranteze, deoarece sunt necesare. căci toate plantele superioare nu au fost încă înfiinţate. Sodiul este absorbit în cantități relativ mari de unele specii din familie. Chenopodiaceae (Chenopodiaceae), în special sfeclă, precum și specii adaptate la condițiile de salinitate, și în acest caz este necesar. Același lucru este valabil și pentru siliciu, care se găsește în cantități deosebit de mari în paiele cerealelor; pentru orez, este un element esențial.
Primele patru elemente - C, H, O, N - se numesc organogeni. Carbonul reprezintă în medie 45% din masa uscată a țesuturilor, oxigen - 42, hidrogen - 6,5 și azot - 1,5 și toate împreună - 95%. Restul de 5% sunt substanțe de cenușă: P, S, K, Ca, Mg, Fe, Al, Si, Na etc. Compoziția minerală a plantelor este de obicei apreciată prin analiza cenușii rămase după arderea materiei organice a plantelor. . Conținutul de elemente minerale (sau oxizi ai acestora) dintr-o plantă este de obicei exprimat ca procent din masa de substanță uscată sau ca procent din masa de cenușă. Substanțele de cenușă enumerate mai sus sunt clasificate ca macronutrienți.
Elementele care sunt prezente în țesuturi în concentrații de 0,001% sau mai puțin din masa uscată a țesuturilor se numesc microelemente. Unele dintre ele joacă un rol important în metabolism (Mn, Cu, Zn, Co, Mo, B, C1).
Conținutul unuia sau altuia din țesuturile plantelor nu este constant și se poate schimba foarte mult sub influența factorilor de mediu. De exemplu, Al, Ni, F și altele se pot acumula în plante la niveluri toxice. Printre plantele superioare, există specii care diferă semnificativ în conținutul în țesuturi a unor astfel de elemente precum Na, așa cum sa menționat deja, și Ca, în legătură cu care grupurile de plante de natriofile, calciofile (majoritatea leguminoase, inclusiv fasole, fasole, trifoi), fobi de calciu (lupin, cu barbă albă, măcriș etc.). Aceste caracteristici specifice se datorează naturii solurilor din locurile de origine și habitatul speciei, un anumit rol genetic fixat pe care aceste elemente îl joacă în metabolismul plantelor.
Frunzele sunt cele mai bogate în elemente minerale, în care cenușa poate fi de la 2 la 15% din masa de substanță uscată. Conținutul minim de cenușă (0,4--1%) a fost găsit în trunchiurile copacilor.
Azotul a fost descoperit în 1772 de chimistul, botanistul și medicul scoțian D. Rutherford ca un gaz care nu sprijină respirația și arderea. Prin urmare, el a fost numit azot, ceea ce înseamnă „non-viață”. Cu toate acestea, azotul face parte din proteine, acizi nucleici și multe substanțe organice vitale. Eliminarea lipsei unor compuși de neînlocuit care conțin azot - aminoacizi, vitamine etc. - este cea mai acută problemă a programelor alimentare ale omenirii.
Azotul este unul dintre cele mai răspândite elemente din natură. Principalele sale forme pe Pământ sunt azotul legat din litosferă și azotul molecular gazos (N 2) din atmosferă, care reprezintă 75,6% din aer în masă. Conform calculelor, rezervele de N 2 din atmosferă sunt estimate la 4 * 10 15 tone.O coloană de aer peste 1 m 2 din suprafața pământului conține 8 tone de azot. Cu toate acestea, azotul molecular ca atare nu este asimilat de plantele superioare și poate fi transformat într-o formă accesibilă acestora doar prin activitatea microorganismelor fixatoare de azot.
Rezervele de azot legat în litosferă sunt, de asemenea, semnificative și sunt estimate la 18 * 10 15 tone.Cu toate acestea, doar o mică parte din azotul litosferic al Pământului este concentrată în sol și doar 0,5 - 2% din rezerva totală în sol. solul este direct disponibil plantelor. 1 hectar de cernoziom arabil conține în medie nu mai mult de 200 kg de azot disponibil plantelor, iar pe podzoli cantitatea sa este de 3-4 ori mai mică. Acest azot este prezent în principal sub formă de ioni NH4+- și NO3-.
microorganisme fixatoare de azot. Microorganismele care efectuează fixarea biologică a azotului pot fi împărțite în două grupe principale: a) fixatori de azot cu viață liberă și b) microorganisme care trăiesc în simbioză cu plantele superioare.
Fixatorii de azot cu viață liberă, heterotrofei, au nevoie de o sursă de carbohidrați de nutriție și, prin urmare, sunt adesea asociați cu microorganisme capabile să descompună celuloza și alte polizaharide. Bacteriile din genurile Azotobacter și Beijerinckia, de regulă, se instalează pe suprafața rădăcinilor plantelor superioare. Astfel de asocieri se explică prin faptul că bacteriile folosesc produse secretate de rădăcini în rizosferă ca sursă de carbon.
Recent, s-a acordat multă atenție cianobacteriilor, în special Tolypothrix avețis. Îmbogățirea câmpurilor de orez cu acestea crește randamentul orezului cu o medie de 20%. În general, valoarea agricolă a fixatorilor de azot cu viață liberă nu este atât de mare. În climatele temperate, fixarea lor anuală de azot este, de regulă, de câteva kilograme de azot la 1 ha, dar în condiții favorabile de sol (de exemplu, o cantitate mare de reziduuri organice), poate ajunge la 20-40 kg N/ha. .
Grupul de fixatori de azot simbiotici include în principal bacterii din genul Rhizobium, care formează noduli pe rădăcinile plantelor leguminoase, precum și unele actinomicete și cianobacterii. În prezent, există aproximativ 190 de specii de plante din familii diferite care pot asimila simbiotic azotul. Printre acestea se numără unii copaci și arbuști: arin, cere, fraier, cătină etc. Nodulii care cresc pe rădăcinile arinului și alte câteva non-leguminoase sunt locuiți de actinomicete din genul Frankia.
Bacteriile nodulare din genul Rhizobium, care trăiesc în simbioză cu plantele leguminoase și fixează în medie 100 până la 400 kg N/ha pe an, prezintă cel mai mare interes pentru agricultură. Dintre leguminoase, lucerna poate acumula până la 500-600 kg N/ha într-un an, trifoiul - 250-300, lupinul - 150, fasole furajeră, mazărea, fasolea - 50-60 kg N/ha. Datorită reziduurilor de cultură și gunoiului verde, aceste plante îmbogățesc în mod semnificativ solul cu azot.
Rezervele de azot din sol pot fi completate în diferite moduri. La cultivarea culturilor agricole, se acordă multă atenție aplicării îngrășămintelor minerale. În condiții naturale, rolul principal revine grupurilor specializate de microorganisme. Aceștia sunt fixatori de azot, precum și bacterii din sol capabile să se mineralizeze și să se transforme în formă de azot organic NH 4 + sau NO 3 - al reziduurilor vegetale și animale care nu sunt disponibile plantelor și azotului din humus, care reprezintă cea mai mare parte a azotului din sol.
Conținutul de azot disponibil plantelor în sol este determinat nu numai de procesele microbiologice de mineralizare a azotului organic și de fixare a azotului, precum și de rata de absorbție a azotului de către plante și de leșierea acestuia din sol, ci și de pierderea acestuia. azotului în procesul de denitrificare efectuat de microorganisme anaerobe capabile să reducă ionul NO3 la N2 gazos. Acest proces este deosebit de intens în soluri umede, inundate, slab aerate, în special în câmpurile de orez.
Astfel, azotul este un element foarte labil care circulă între atmosferă, sol și organismele vii.
1.3 Fosfor
Fosforul, ca și azotul, este un nutrient esențial pentru plante. Este absorbit de ei sub formă de oxid superior PO 4 3- și nu se modifică, fiind inclus în compușii organici. În țesuturile plantelor, concentrația de fosfor este de 0,2--1,3% din masa uscată a plantei.
Forme de compuși ai fosforului disponibile plantelor
Rezervele de fosfor din stratul arabil al solului sunt relativ mici, aproximativ 2,3 - 4,4 t/ha (din punct de vedere al P 2 O 5). Din această cantitate, 2/3 cade pe sărurile minerale ale acidului ortofosforic (H 3 PO 4), iar 1/3 - pe compușii organici care conțin fosfor (reziduuri organice, humus, fitat etc.). Fitații reprezintă până la jumătate din fosforul organic din sol. Majoritatea compușilor de fosfor sunt ușor solubili în soluția din sol. Aceasta, pe de o parte, reduce pierderea de fosfor din sol din cauza levigarii, dar, pe de altă parte, limitează posibilitățile de utilizare a acestuia de către plante.
Principala sursă naturală de fosfor din stratul arabil este degradarea rocilor formatoare de sol, unde se găsește în principal sub formă de apatite 3Ca 3 (P0 4) 2 * CaF 2 etc. Săruri fosforice trisubstituite de calciu și magneziu iar sărurile sesquioxizilor de fier și aluminiu (FeP0 4, AIPO 4 în solurile acide) sunt slab solubile și greu accesibile plantelor. Sărurile disubstituite și mai ales monosubstituite de calciu și magneziu, în special sărurile de cationi monovalenți și acid fosforic liber, sunt solubile în apă și sunt utilizate de plante ca sursă principală de fosfor în soluția solului. Plantele sunt, de asemenea, capabile să asimileze unele forme organice de fosfor (fosfați de zahăr, fitină). Concentrația de fosfor în soluția de sol este scăzută (0,1 - 1 mg / l). Fosforul din reziduurile organice și humus este mineralizat de microorganismele din sol și cea mai mare parte este transformat în săruri puțin solubile. Plantele obțin fosfor din ele, făcându-l mai mobil. Acest lucru se realizează datorită eliberării de acizi organici de către rădăcini, care chelează cationii divalenți și acidifică rizosfera, facilitând tranziția HPO 4 3-> HPO 4 2-> HP0 4 - . Unele culturi absorb bine fosfații puțin solubili (lupin, hrișcă, mazăre). Această capacitate la plante crește odată cu vârsta.
Participarea fosforului în metabolism
În țesuturile plantelor, fosforul este prezent sub formă organică și sub formă de acid fosforic și sărurile sale. Face parte din proteine (fosfoproteine), acizi nucleici, fosfolipide, esteri de fosfat de zahăr, nucleotide implicate în metabolismul energetic (ATP, NAD+ etc.), vitamine și mulți alți compuși.
Fosforul joacă un rol deosebit de important în energia celulei, deoarece este sub formă de legături eterice de înaltă energie ale fosforului (C--O ~ P) sau legături pirofosfat în nucleozide di-, nucleozide trifosfați și polifosfați. depozitat într-o celulă vie. Aceste legături au o energie liberă standard ridicată de hidroliză (de exemplu, 14 kJ/mol pentru glucoză-6-fosfat și AMP, 30,5 pentru ADP și ATP și 62 kJ/mol pentru fosfoenolpiruvat). Acesta este un mod atât de universal de stocare și utilizare a energiei, încât aproape toate căile metabolice implică anumiți esteri fosforici și (sau) nucleotide, iar starea sistemului de nucleotide adeninei (încărcare energetică) este un mecanism important pentru controlul respirației.
Sub formă de diester stabil, fosfatul este o parte integrantă a structurii acizilor nucleici și fosfolipidelor. În acizii nucleici, fosforul formează punți între nucleozide, unindu-le într-un lanț uriaș. Fosfatul face fosfolipidul hidrofil, în timp ce restul moleculei este lipofil. Prin urmare, la limita de fază în membrane, moleculele de fosfolipide sunt orientate polar, cu capetele lor fosfat în exterior, iar miezul lipofil al moleculei este ținut ferm în stratul dublu lipidic, stabilizând membrana.
O altă funcție unică a fosforului este participarea sa la fosforilarea proteinelor celulare de către protein kinaze. Acest mecanism controlează multe procese metabolice, deoarece includerea fosfatului într-o moleculă de proteină duce la o redistribuire a sarcinilor electrice în ea și, ca urmare, la o modificare a structurii și funcției sale. Fosforilarea proteinelor reglează procese precum sinteza ARN-ului și proteinelor, diviziunea celulară, diferențierea celulară și multe altele.
Principala formă de rezervă a fosforului din plante este fitina - sarea de calciu-magneziu a acidului fosforic inozitol (inozitol hexafosfat):
În semințe se acumulează cantități semnificative de fitină (0,5-2% din greutatea uscată), reprezentând până la 50% din fosforul total din acestea.
Mișcarea radială a fosforului în zona de absorbție a rădăcinii la xilem are loc de-a lungul simplastului, iar concentrația sa în celulele rădăcinii este de zeci până la sute de ori mai mare decât concentrația de fosfat din soluția de sol. Transportul prin xilem se realizează în principal sau în întregime sub formă de fosfat anorganic; sub această formă ajunge la frunze și zonele de creștere. Fosforul, ca și azotul, este ușor redistribuit între organe. Din celulele frunzelor, intră în tuburile sită și este transportată de-a lungul floemului către alte părți ale plantei, în special către conurile de creștere și fructele în curs de dezvoltare. Un flux similar de fosfor are loc din frunzele îmbătrânite.
Sulful este unul dintre principalii nutrienți necesari vieții plantelor. Intră în ele în principal sub formă de sulfat. Conținutul său în țesuturile plantelor este relativ scăzut și se ridică la 0,2--1,0% raportat la greutatea uscată. Nevoia de sulf este mare în plantele bogate în proteine, precum leguminoasele (lucernă, trifoi), dar este deosebit de pronunțată la reprezentanții familiei cruciferelor, care sintetizează uleiuri de muștar care conțin sulf în cantități mari.
Sulful se găsește în sol în forme anorganice și organice. În majoritatea solurilor predomină sulful organic din resturi vegetale și animale, iar în solurile turboase poate reprezenta până la 100% din totalul sulfului. Principala formă anorganică a sulfului din sol este sulfatul, care poate fi sub formă de săruri CaSO 4 , MgSO 4 , Na 2 SO 4 în soluția de sol în formă ionică sau adsorbită pe coloizii din sol. În solurile saline Na 2 SO 4, conținutul de sulfat poate ajunge la 60% din masa solului. În solurile inundate, sulful este în formă redusă sub formă de FeS, FeS 2 sau H 2 S. Conținutul total de sulf în solurile din zonele climatice temperate este în medie de 0,005 - 0,040%.
Plantele absorb sulful în principal sub formă de sulfat. Transferul transmembranar al sulfatului se realizează în co-transport cu H + sau în schimbul ionilor HCO 3 -. Compușii anorganici cu sulf mai puțin oxidați (SO2) sau mai redusi (H3S) sunt toxici pentru plante. Plantele și compușii organici (aminoacizi) care conțin sulf redus sunt foarte prost percepute.
Sulful se găsește în plante în două forme principale - oxidat (sub formă de sulfat anorganic) și redus. Conținutul absolut și raportul formelor oxidate și reduse de sulf în organele plantelor depind atât de activitatea proceselor de reducere și asimilare a sulfatului care au loc în acestea, cât și de concentrația de SO 4 2- în mediul nutritiv.
O parte din sulful absorbit de plantă este reținută în bazinul de sulfat de rădăcină, eventual sub formă de CaSO 4 sau sulfat metabolic, care este nou format ca urmare a oxidării secundare a sulfului redus. Partea principală a sulfatului se deplasează de la rădăcini la vasele de xilem și este transferată cu curentul de transpirație către organele tinere în creștere, unde este inclusă intens în metabolism și își pierde mobilitatea.
Din frunze, sulfatul și formele reduse de sulf (aminoacizi care conțin sulf, glutation) se pot deplasa de-a lungul floemului atât acropet, cât și bazipet, către părțile în creștere ale plantelor și organele de depozitare. În semințe, sulful este preponderent într-o formă organică, iar în procesul de germinare a acestora se transformă parțial într-o formă oxidată. Reducerea sulfatului și sinteza aminoacizilor și proteinelor care conțin sulf se observă în timpul maturării semințelor.
Proporția de sulfat în soldul total de sulf din țesuturi poate varia de la 10 la 50% sau mai mult. Este minim în frunzele tinere și crește brusc odată cu îmbătrânirea lor datorită degradării crescute a proteinelor care conțin sulf.
Sulful face parte din cei mai importanți aminoacizi - cisteina și metionina, care se găsesc în plante, atât sub formă liberă, cât și ca parte a proteinelor. Metionina este unul dintre cei 10 aminoacizi esențiali și are proprietăți unice datorită grupării sale de sulf și metil.
Una dintre funcțiile principale ale sulfului în proteine și polipeptide este participarea grupărilor SH la formarea legăturilor covalente, de hidrogen și mercaptide care susțin structura tridimensională a proteinei.
Sulful este, de asemenea, o parte a celor mai importanți compuși biologici - coenzima A și vitaminele (acid lipoic, biotină, tiamină) și sub forma acestor compuși participă la reacțiile enzimatice ale celulei.
Potasiul este unul dintre cele mai esențiale elemente ale nutriției minerale a plantelor. Conținutul său în țesuturi este în medie de 0,5 - 1,2% pe baza greutății uscate. Multă vreme, cenușa a servit ca sursă principală de potasiu, ceea ce se reflectă în numele elementului (potasiul provine de la cuvântul potasi - cenușă de creuzet). Conținutul de potasiu din celulă este de 100-1000 de ori mai mare decât nivelul său din mediul extern. Este mult mai mult în țesuturi decât alți cationi.
Rezervele de potasiu din sol sunt de 8 până la 40 de ori mai mari decât conținutul de fosfor, iar azotul de 5 până la 50 de ori. În sol, potasiul poate fi sub următoarele forme: ca parte a rețelei cristaline a mineralelor, în stare de schimb și neschimb în particule coloidale, ca parte a reziduurilor de cultură și microorganismelor, sub formă de săruri minerale ale solului soluţie.
Cea mai bună sursă de nutriție sunt sărurile de potasiu solubile (0,5 - 2% din rezervele brute din sol). Pe măsură ce se consumă formele mobile de potasiu, rezervele sale din sol pot fi reînnoite în detrimentul formelor interschimbabile, iar când acestea din urmă scad, în detrimentul formelor fixe de potasiu neschimbabile. Uscarea și umezirea alternativă a solului, precum și activitatea sistemului radicular al plantelor și microorganismelor, contribuie la tranziția potasiului în forme accesibile.
La plante, potasiul este concentrat în cea mai mare cantitate în țesuturile tinere, în creștere, caracterizate printr-un nivel ridicat de metabolism: meristeme, cambium, frunze tinere, lăstari, muguri. În celule, potasiul este prezent în principal sub formă ionică; nu face parte din compușii organici, are mobilitate mare și, prin urmare, este ușor de reutilizat. Mișcarea potasiului de la frunzele bătrâne la cele tinere este facilitată de sodiu, care îl poate înlocui în țesuturile plantelor care au încetat să crească.
În celulele vegetale, aproximativ 80% din potasiu este conținut în vacuole. El constituie cea mai mare parte a cationilor sevei celulare. Prin urmare, potasiul poate fi spălat din plante de ploi, în special din frunzele bătrâne. În timpul înfometării cu potasiu, structura lamelară-granulară a cloroplastelor este perturbată, iar structurile membranare ale mitocondriilor sunt dezorganizate. Până la 20% din potasiul celulei este adsorbit pe coloizii citoplasmei. La lumină, puterea de legătură a potasiului cu coloizii este mai mare decât la întuneric. Noaptea poate fi observată chiar și eliberarea de potasiu prin sistemul radicular al plantelor.
Potasiul contribuie la mentinerea starii de hidratare a coloizilor citoplasmei, reglandu-i capacitatea de retinere a apei. O creștere a hidratării proteinelor și a capacității de reținere a apei a citoplasmei crește rezistența plantelor la secetă și îngheț.
Potasiul este esențial pentru absorbția și transportul apei în întreaga plantă. Calculele arată că munca „motorului inferior”, adică presiunea rădăcinii, este de 3/4 din cauza prezenței ionilor de potasiu în seva. Potasiul joacă un rol important în procesul de deschidere și închidere a stomatelor. În lumină, în vacuolele celulelor de gardă ale stomatelor, concentrația ionilor de potasiu crește brusc (de 4-5 ori), ceea ce duce la o intrare rapidă a apei, o creștere a turgenței și deschiderea fisurii stomatice. Pe întuneric, potasiul începe să părăsească celulele de gardă, presiunea turgenței din ele scade și stomatele se închid.
Potasiul este preluat de plante ca cation și formează doar legături slabe cu diverși compuși din celulă. Acesta este probabil motivul pentru care potasiul este cel care creează asimetria ionică și diferența de potențiale electrice dintre celulă și mediu (potențial de membrană).
Potasiul este unul dintre cationii - activatori ai sistemelor enzimatice. În prezent, se știe că peste 60 de enzime sunt activate de potasiu cu diferite grade de specificitate. Este necesar pentru încorporarea fosfatului în compuși organici, reacții de transfer ale grupărilor fosfat, pentru sinteza proteinelor și polizaharidelor și este implicat în sinteza riboflavinei, o componentă a tuturor flavindehidrogenazelor. Sub influența potasiului, crește acumularea de amidon în tuberculii de cartofi, zaharoza în sfecla de zahăr, monozaharidele din fructe și legume, celuloza, hemicelulozele și substanțele pectinice în peretele celular al plantelor. Ca urmare, rezistența paielor de cereale la adăpost crește, iar calitatea fibrei se îmbunătățește la in și cânepă. Aprovizionarea suficientă cu potasiu a plantelor crește rezistența acestora la bolile fungice și bacteriene.
1,6 calciu
Conținutul total de calciu la diferite specii de plante este de 5-30 mg la 1 g de greutate uscată. Plantele în raport cu calciul sunt împărțite în trei grupe: calciofile, calciofobe și specii neutre. O mulțime de calciu conține leguminoase, hrișcă, floarea soarelui, cartofi, varză, cânepă, mult mai puțin - cereale, in, sfeclă de zahăr. În țesuturile plantelor dicotiledone, acest element, de regulă, este mai mare decât la monocotiledone.
Calciul se acumulează în organele și țesuturile vechi. Acest lucru se datorează faptului că transportul acestuia se realizează prin xilem și reutilizarea este dificilă. Odată cu îmbătrânirea celulelor sau scăderea activității lor fiziologice, calciul din citoplasmă se deplasează în vacuole și se depune sub formă de săruri insolubile ale acizilor oxalic, citric și alți acizi. Incluziunile cristaline rezultate împiedică mobilitatea și reutilizarea acestui cation.
La majoritatea plantelor cultivate, calciul se acumulează în organele vegetative. În sistemul radicular, conținutul său este mai scăzut decât în partea aeriană. În semințe, calciul este prezent în principal sub formă de sare a acidului inozitol-fosforic (fitina).
Calciul îndeplinește o varietate de funcții în metabolismul celulelor și al corpului în ansamblu. Ele sunt asociate cu influența acesteia asupra structurii membranelor, a fluxurilor de ioni prin acestea și a fenomenelor bioelectrice, asupra rearanjarii citoscheletului, a proceselor de polarizare a celulelor și țesuturilor etc.
Calciul activează o serie de sisteme enzimatice celulare: dehidrogenaze (glutamat dehidrogenază, malat dehidrogenază, glucozo-6-fosfat dehidrogenază, izocitrat dehidrogenază dependentă de NADP), b amilază, adenilat și arginin kinaze, lipaze, fosfataze. În acest caz, calciul poate promova agregarea subunităților proteice, poate servi ca o punte între enzimă și substrat și poate afecta starea centrului alosteric al enzimei. Excesul de calciu sub formă ionică inhibă fosforilarea oxidativă și fotofosforilarea.
Un rol important revine ionilor de Ca 2 + în stabilizarea membranelor. Interacționând cu grupurile de fosfolipide încărcate negativ, stabilizează membrana și îi reduce permeabilitatea pasivă. Cu lipsa de calciu, permeabilitatea membranelor crește, apar rupturi și fragmentare ale acestora, iar procesele de transport membranar sunt perturbate.
Este important de menționat că aproape întreaga capacitate de schimb de cationi a suprafeței rădăcinii este ocupată de calciu și parțial de H+. Aceasta indică implicarea calciului în mecanismele primare de intrare a ionilor în celulele radiculare. Limitând intrarea altor ioni în plante, calciul ajută la eliminarea toxicității concentrațiilor în exces de ioni de amoniu, aluminiu, mangan și fier, crește rezistența plantelor la salinitate și reduce aciditatea solului. Este calciul care acționează cel mai adesea ca un ion de echilibru atunci când creează un echilibru fiziologic al compoziției ionice a mediului, deoarece conținutul său în sol este destul de mare.
Cele mai multe tipuri de sol sunt bogate în calciu, iar înfometarea pronunțată de calciu este rară, de exemplu, cu aciditate puternică sau salinitate a solurilor, pe turbării, cu o încălcare a dezvoltării sistemului radicular, în condiții meteorologice nefavorabile.
1,7 magneziu
În ceea ce privește conținutul din plante, magneziul ocupă locul patru după potasiu, azot și calciu. La plantele superioare, conținutul său mediu în ceea ce privește greutatea uscată este de 0,02 - 3,1%, la alge 3,0 - 3,5%. Mai ales o mulțime din plantele de zi scurtă - porumb, mei, sorg, cânepă, precum și cartofi, sfeclă, tutun și leguminoase. 1 kg de frunze proaspete conține 300 - 800 mg de magneziu, din care 30 - 80 mg (adică 1/10 parte) fac parte din clorofilă. Există mai ales mult magneziu în celulele tinere și țesuturile în creștere, precum și în organele generatoare și țesuturile de depozitare. În cariopse, magneziul se acumulează în embrion, unde nivelul său este de câteva ori mai mare decât conținutul din endosperm și coajă (pentru porumb, respectiv, 1,6, 0,04 și 0,19% pe greutatea uscată).
Acumularea de magneziu în țesuturile tinere este facilitată de mobilitatea sa relativ mare în plante, ceea ce duce la utilizarea secundară (reutilizare) din țesuturile îmbătrânite. Totuși, gradul de reutilizare a magneziului este mult mai scăzut decât cel al azotului, fosforului și potasiului. Mobilitatea ușoară a magneziului se explică prin faptul că aproximativ 70% din acest cation din plantă este asociat cu anioni ai acizilor organici și anorganici. Mișcarea magneziului se realizează atât în xilem, cât și în floem. O parte din magneziu formează compuși insolubili care nu sunt capabili să se deplaseze prin plantă (oxalat, pectat), cealaltă parte este legată de compuși macromoleculari. În semințe (embrion, coajă), cea mai mare parte a magneziului se află în compoziția fitinei.
Și, în sfârșit, aproximativ 10-12% din magneziu face parte din clorofilă. Această ultimă funcție a magneziului este unică: niciun alt element nu o poate înlocui în clorofilă. Magneziul este necesar pentru sinteza protoporfirinei IX, precursorul direct al clorofilei.
În lumină, ionii de magneziu sunt eliberați din cavitatea tilacoizilor în stroma cloroplastei. O creștere a concentrației de magneziu în stromă activează RDF-carboxilaza și alte enzime. Se presupune că o creștere a concentrației de Mg 2 + (până la 5 mmol/l) în stromă duce la o creștere a afinității RDP carboxilază pentru CO 2 și la activarea reducerii CO 2 . Magneziul poate afecta direct conformația enzimei, precum și să ofere condiții optime pentru funcționarea acesteia prin afectarea pH-ului citoplasmei ca contraion de protoni. Ionii de potasiu pot acționa similar. Magneziul activează o serie de reacții de transfer de electroni în timpul fotofosforilării: reducerea NADP+, viteza reacției Hill, este necesar pentru transferul de electroni de la PS II la PS I.
Acțiunea magneziului asupra altor zone ale metabolismului este cel mai adesea asociată cu capacitatea sa de a regla activitatea enzimelor, iar semnificația sa pentru o serie de enzime este unică. Doar manganul poate înlocui magneziul în unele procese. Cu toate acestea, în majoritatea cazurilor, activarea enzimelor de către magneziu (la concentrație optimă) este mai mare decât de către mangan.
Magneziul este esențial pentru multe enzime de glicoliză și pentru ciclul Krebs. În mitocondrii, cu deficiența sa, se observă o scădere a numărului, o încălcare a formei și, în cele din urmă, dispariția cristei. Nouă dintre cele douăsprezece reacții de glicoliză necesită participarea activatorilor metalici, iar șase dintre ele sunt activate de magneziu.
Magneziul intensifică sinteza uleiurilor esențiale, cauciucului, vitaminelor A și C. Se presupune că, formând un compus complex cu acidul ascorbic, întârzie oxidarea acestuia. Mg2+ este necesar pentru formarea ribozomilor și polizomilor, pentru activarea aminoacizilor și sinteza proteinelor și este utilizat pentru toate procesele la o concentrație de cel puțin 0,5 mmol/l. Activează ADN și ARN polimeraze, participă la formarea unei anumite structuri spațiale a acizilor nucleici.
Odată cu creșterea gradului de aprovizionare cu magneziu în plante, crește conținutul de compuși ai fosforului în forme organice și anorganice. Acest efect este probabil asociat cu rolul magneziului în activarea enzimelor implicate în metabolismul fosforului.
Plantele se confruntă cu o lipsă de magneziu în principal pe solurile nisipoase. Solurile podzolice sunt sărace în magneziu și calciu, serozem-urile sunt bogate; cernoziomurile ocupă o poziţie intermediară. Magneziu solubil în apă și schimbabil în sol 3--10%. Complexul de absorbție a solului conține cei mai mulți ioni de calciu, magneziul se află pe locul doi. Plantele se confruntă cu o lipsă de magneziu în cazurile în care acesta conține mai puțin de 2 mg la 100 g de sol. Odată cu scăderea pH-ului soluției din sol, magneziul pătrunde în plante în cantități mai mici.
CAPITOLUL 2. MATERIALE ȘI METODE DE CERCETARE
2.1 Metode de determinare a mineralelor
Determinarea conținutului oricărui element chimic dintr-o plantă include, ca procedură obligatorie premergătoare determinării în sine, etapa de descompunere (digestie) a probei.
În practica analizei biochimice, se folosesc în principal două metode - cenușa uscată și umedă. În ambele cazuri, procedura asigură mineralizarea tuturor elementelor, adică conversia lor într-o formă care este solubilă într-unul sau altul solvent anorganic.
Cenușa umedă este metoda principală de descompunere a compușilor organici ai azotului și fosforului și, în unele cazuri, este mai fiabilă în determinarea multor alte elemente. La determinarea borului, se poate folosi numai cenușă uscată, deoarece majoritatea compușilor de bor se volatilizează cu apă și vapori acizi.
Metoda de incinsare uscată este aplicabilă analizei conținutului aproape tuturor macro și microelementelor din materialul biologic. De obicei, cenușarea uscată a probelor de plante se realizează într-un cuptor electric cu mufă în creuzete (sau cupe) de porțelan, cuarț sau metal, la o temperatură care nu depășește 450-500 ° C. Crezetele de cuarț sunt cele mai bune, dar creuzetele din sticlă refractară sau porțelan sunt de obicei folosite. Unele studii speciale pot necesita creuzete de platină. Temperatura scazuta in timpul arderii si alegerea corecta a materialului creuzetului fac posibila evitarea pierderilor prin volatilizare si a pierderilor datorate formarii de oxizi ai elementului aflat in determinare care sunt slab solubili in acid clorhidric. Oxizii pot fi formați prin reacția cu materialul din care sunt fabricate creuzetele.
2.2 Analiza microchimică a cenușii
Materiale si utilaje: cenusa obtinuta prin arderea frunzelor, semintelor, lemnului; Soluții 10% HCI și NH3, soluții 1% din următoarele săruri într-un picurător: Na 2 HCO 3 , NaHC 4 H 4 O 6 , K 4 , (NH 4 ) 2 MoO 4 în 1% HNO 3 , 1% H 2 SO soluţie 4 ; eprubete, pâlnii de sticlă cu diametrul de 4-5 cm, spatule metalice sau spatule pentru ochi, lamele de sticlă, baghete de sticlă, șervețele sau bucăți de hârtie de filtru, filtre de hârtie, șaibe sau baloane cu apă distilată, căni pentru spălarea apei.
Informatie scurta:
Când țesutul este ars, elementele organogenice (C; H; O; N) se evaporă sub formă de compuși gazoși, iar partea incombustibilă rămâne - cenușă. Conținutul său în diferite organe este diferit: într-o frunză - până la 10-15%, în semințe - aproximativ 3%, în lemn - aproximativ 1%. Cea mai mare parte a cenușii se află în țesuturile vii, care funcționează activ, de exemplu, în mezofila frunzei. Celulele sale conțin clorofilă și multe enzime, care includ elemente precum magneziu, fier, cupru etc. Datorită activității metabolice ridicate a țesuturilor vii, acestea conțin și o cantitate semnificativă de potasiu, fosfor și alte elemente. Conținutul de cenușă depinde de compoziția solului pe care crește planta și de vârsta și natura sa biologică. Organele plantelor diferă nu numai în ceea ce privește compoziția cantitativă, ci și calitativă a cenușii.
Metoda microchimică face posibilă detectarea unui număr de elemente în cenușa plantelor. Metoda se bazează pe capacitatea unor reactivi, atunci când interacționează cu elementele de cenușă, de a da compuși care diferă într-o anumită culoare sau formă a cristalelor.
Proces de lucru
O parte din materialul uscat (așchii de lemn, frunze și semințe zdrobite) se pune într-un creuzet, se adaugă puțin alcool și se dă foc. Repetați procedura de 2-3 ori. Apoi transferați creuzetul într-o sobă electrică și aprindeți până când materialul carbonizat capătă o culoare cenușiu. Cărbunele rămas trebuie ars prin introducerea creuzetului într-un cuptor cu mufă timp de 20 de minute.
Pentru a detecta Ca, Mg, P și Fe, este necesar să adăugați o porțiune de cenușă în eprubetă cu o spatulă de ochi de sticlă, să o umpleți cu 4 ml de HCl 10% și să o agitați de câteva ori pentru o mai bună dizolvare. Pentru a detecta potasiul, aceeași cantitate de cenușă trebuie dizolvată în 4 ml de apă distilată și filtrată într-o eprubetă curată printr-un filtru mic de hârtie. Apoi, cu o baghetă de sticlă, se pune o picătură mică de extract de cenușă pe o lamă de sticlă curată, apoi, la o distanță de 10 mm, o picătură de reactiv și se folosește un bețișor pentru a conecta două picături cu un jumper. (Fiecare reactiv se aplică cu o pipetă separată). În punctul de contact al soluțiilor, va avea loc cristalizarea produselor de reacție (amestecarea a două picături este nedorită, deoarece se formează cristale mici atipice din cauza cristalizării rapide; în plus, când picătura se usucă, se pot forma cristale ale sărurilor inițiale) .
După aceea, îndepărtați picăturile de soluții rămase din sticlă cu bucăți de hârtie de filtru și examinați cristalele la microscop fără lamelă. După fiecare reacție, tija de sticlă trebuie clătită cu apă și uscată cu hârtie de filtru.
Pentru detectarea potasiului se folosește tartrat de sodiu acid 1%. În urma reacţiei cu un extract de cenuşă se formează cristale de tartrat acid de potasiu KHC 4 H 4 O 6, având forma unor prisme mari. Extractul de potasiu în apă trebuie mai întâi neutralizat, deoarece produsul de reacție este solubil într-un mediu acid și alcalin. Reacția decurge conform ecuației:
NaHC 4 H 4 O 6 + K + > KNS 4 H 4 O 6 v + Na + .
Detectarea calciului se realizează cu acid sulfuric 1%, reacția se desfășoară conform ecuației:
CaCI2 + H2S04 > CaS04 v + 2HCI.
Ca rezultat, ghipsul se formează sub formă de cristale separate sau grupate în formă de ac.
Când este detectat magneziu, o picătură de soluție de amoniac 10% este adăugată mai întâi la o picătură de extract de cenușă și conectată printr-o punte cu o picătură de soluție de fosfat de sodiu 1%. Reacția decurge conform ecuației:
MgCI2 + NH3 + Na2HP04 > NH4MgP04v + 2NaCI.
Sarea fosfor-amoniac-magnezie se formează sub formă de cristale plate incolore sub formă de dreptunghiuri, aripi, capace.
Detectarea fosforului se realizează folosind molibdat de amoniu 1% în acid azotic. Reacția decurge conform ecuației:
H3PO4 + 12 (NH4)2MoO4+21HNO3>(NH4)3PO4*12MoO3v + 21NH4NO3 + 12H2O.
Amoniacul fosfor-molibden se formează sub formă de bucăți mici de culoare galben-verde.
Pentru a detecta fierul, se toarnă o cantitate egală de extract de cenușă din diferite organe (1-2 ml) în două eprubete, se adaugă o cantitate egală de sare de sânge galbenă 1% până când apare o culoare albastră. Se formează albastrul prusac:
4FeCl 3 + 3K 4 > Fe 4 3 + 12KCl.
CAPITOLUL 3. REZULTATELE CERCETĂRII ȘI ANALIZA LOR
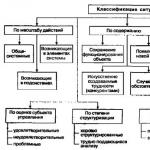
3.1 Simptome ale deficienței minerale
Lipsa mineralelor determină modificări ale proceselor biochimice și fiziologice, în urma cărora se observă adesea modificări morfologice, sau simptome vizibile.
Uneori, din cauza deficienței, creșterea este suprimată înainte de apariția altor simptome.
Simptome vizibile de deficiență. Cel mai semnificativ rezultat al lipsei de minerale este creșterea redusă. Totuși, efectul cel mai vizibil este îngălbenirea frunzelor, cauzată de scăderea biosintezei clorofilei. Frunzele par a fi deosebit de susceptibile la deficiență. Cu lipsa mineralelor, dimensiunea acestora scade, forma sau structura se schimba, culoarea devine palida, iar uneori se formeaza chiar zone moarte la varfuri, margini sau intre venele principale. În unele cazuri, frunzele sunt adunate în ciorchini sau rozete, iar acele de pin uneori nu reușesc să se separe și se formează „ace îmbinate”. Un simptom comun al unui anumit tip de deficiență minerală la plantele erbacee este suprimarea creșterii tulpinii și reducerea creșterii limbei frunzelor, rezultând rozete de frunze mici, adesea cu o rețea de pete clorotice. Simptomele vizibile ale deficienței diferitelor elemente sunt atât de caracteristice încât observatorii experimentați pot identifica deficiența după aspectul frunzelor.
Uneori, cu lipsa de minerale, copacii formează cantități în exces de gumă. Acest fenomen se numește homosis. Secreția de rășină în jurul mugurilor este caracteristică pinilor cu deficit de zinc din Australia. Guma se găsește și pe scoarța pomilor fructiferi care suferă de moartea cauzată de lipsa de cupru. Deficiența severă provoacă adesea moartea frunzelor, lăstarilor și a altor părți, adică se dezvoltă simptomele descrise ca moarte. Moartea lăstarilor cauzată de lipsa cuprului a fost observată în multe păduri și pomi fructiferi. Când lăstarii apicali mor, merii care suferă de deficiență de cupru capătă un aspect stufșat, piper. Lipsa borului determină uscarea punctelor apicale de creștere și în cele din urmă moartea cambiumului la citrice și pini, moartea floemului și degradarea fiziologică a fructelor la alte specii. Lipsa unui element contribuie uneori la apariția mai multor simptome diferite, de exemplu, deficiența de bor la meri provoacă deformarea și fragilitatea frunzelor, necroza floemului, deteriorarea scoarței și fructelor.
cloroza. Cel mai frecvent simptom observat cu lipsa unei varietăți mari de elemente este cloroza, rezultată dintr-o încălcare a biosintezei clorofilei. Natura, gradul și severitatea clorozei la frunzele tinere și bătrâne depind de tipul de plantă, elementul și gradul de deficiență. Cel mai adesea, cloroza este asociată cu o lipsă de azot, dar poate fi cauzată și de o deficiență de fier, mangan, magneziu, potasiu și alte elemente. În plus, cloroza poate fi cauzată nu numai de o deficiență de minerale, ci și de o varietate de alți factori negativi de mediu, inclusiv prea multă sau prea puțină apă, temperaturi nefavorabile, substanțe toxice (cum ar fi dioxidul de sulf) și un exces de minerale. Cloroza poate fi cauzată și de factori genetici care provoacă apariția unor plante variate colorate: de la albinos, complet lipsiți de clorofilă, până la răsaduri verzui sau răsad cu diverse dungi și pete de frunze.
Pe baza numeroșilor factori care provoacă cloroza, se poate concluziona că aceasta apare atât ca urmare a unei tulburări metabolice generale, cât și a influenței specifice a elementelor individuale.
Unul dintre cele mai frecvente tipuri de cloroză care provoacă cele mai multe perturbări în dezvoltarea plantelor este cea întâlnită într-un număr mare de pomi fructiferi, ornamentali și forestieri care cresc pe soluri alcaline și calcaroase. De obicei este cauzată de inaccesibilitatea fierului la valori ridicate ale pH-ului, dar uneori cauza este o deficiență de mangan.
În cloroza la angiosperme, nervurile mijlocii și mai mici ale frunzelor rămân verzi, iar zonele dintre nervuri devin verde pal, galben sau chiar alb. De obicei, cele mai tinere frunze sunt cele mai afectate de cloroză. La conifere, acele tinere devin verde pal sau galbene, iar cu o deficiență mare, acele pot deveni maro și se desprind.
Cloroza cauzată de deficitul de fier poate fi eliminată parțial sau complet prin scăderea pH-ului solului.
3.2 Efectul fiziologic al deficitului de minerale
Efectele morfologice vizibile sau simptomele deficienței minerale sunt rezultatul modificărilor diferitelor procese biochimice sau fiziologice interne. Cu toate acestea, din cauza relațiilor complexe dintre ele, poate fi dificil să se determine modul în care deficiența unui anumit element provoacă efectele observate. De exemplu, lipsa de azot poate inhiba creșterea din cauza unei aprovizionări mai slabe de azot la procesele de biosinteză a noii protoplasme. Dar, în același timp, rata de sinteză a enzimelor și a clorofilei scade, iar suprafața fotosintetică scade. Acest lucru determină o slăbire a fotosintezei, ceea ce înrăutățește aprovizionarea proceselor de creștere cu carbohidrați. Ca rezultat, este posibilă o scădere suplimentară a ratei de absorbție a azotului și a mineralelor. Un element îndeplinește adesea mai multe funcții într-o plantă, așa că nu este ușor de determinat care funcție sau combinație de funcții este perturbată provocând simptome vizibile. Manganul, de exemplu, pe lângă activarea anumitor sisteme enzimatice, este necesar și pentru sinteza. Clorofilă. Deficiența acestuia provoacă unele tulburări funcționale. Lipsa de azot duce de obicei la o scădere marcată a fotosintezei, dar efectul lipsei altor elemente nu este atât de clar.
Lipsa mineralelor reduce atât biosinteza carbohidraților, cât și mișcarea acestora către țesuturile în creștere. Deficiența afectează adesea fotosinteza și respirația în mod diferit. De exemplu, o deficiență semnificativă de potasiu încetinește fotosinteza și crește respirația, reducând astfel cantitatea de carbohidrați care pot fi utilizați pentru creștere. Uneori, mișcarea carbohidraților este de asemenea suprimată. Acest efect este pronunțat la arborii cu deficit de bor cu necroză a floemului. Ca urmare a scăderii cantității de carbohidrați disponibili, rata de creștere a țesuturilor într-o parte a copacului este redusă, dar, în același timp, poate apărea o acumulare de carbohidrați în altă parte. Uneori, datorită conținutului scăzut de carbohidrați de rezervă, formarea semințelor este redusă. Aplicarea abundentă a îngrășământului cu azot a dus la o creștere semnificativă a procesului de formare a semințelor la fag și arțar de zahăr, a crescut procentul semințelor sănătoase și greutatea uscată a semințelor de arțar. Formarea conurilor și a semințelor la pinul tânăr de tămâie a crescut, de asemenea, brusc după fertilizare. Dacă pomii nu au deficit de minerale, aplicarea unor cantități mari de îngrășăminte cu azot poate reduce formarea fructelor și semințelor datorită stimulării creșterii vegetative.
3.3 Excesul de minerale
Solurile de pădure au rareori un exces de nutrienți minerali, dar fertilizarea abundentă a livezilor și a pepinierelor duce uneori la concentrații de sare suficient de mari pentru a provoca rău. Există, de asemenea, suprafețe mari de terenuri aride unde majoritatea speciilor de plante nu pot exista din cauza conținutului ridicat de sare. Irigarea cu apă care conține multă sare provoacă, de asemenea, daune. Acest lucru se datorează unei creșteri a presiunii osmotice, modificări nefavorabile ale pH-ului pentru plante, dezechilibru al diferiților ioni sau ca urmare a unei combinații a acestor factori.
Presiunea osmotică crescută a soluției de sol reduce absorbția de apă, crește deficitul de apă în frunze și duce la deteriorarea țesuturilor prin uscare în zilele în care vântul și temperaturile ridicate provoacă transpirație puternică. Cu o deshidratare mai lungă și mai profundă, stomatele se închid și ele, prevenind fotosinteza. Concentrațiile mari de sare în sol pot provoca deteriorarea rădăcinilor prin plasmoliză, în special în solurile nisipoase, care interferează cu activitatea sintetică a rădăcinilor. Uneori, frunzele sunt deteriorate ca urmare a concentrațiilor mari de îngrășăminte lichide care le sunt aplicate.
Efectul nociv al fertilizării excesive depinde de tipul de plantă, de tipul de îngrășământ folosit și de momentul aplicării.
Suprafertilizarea pomilor fructiferi și ornamentali prelungește uneori sezonul de creștere într-o asemenea măsură încât copacii și arbuștii nu au timp să dobândească rezistență la frig înainte de îngheț. Fertilizarea excesivă stimulează uneori formarea unui număr mare de ramuri, flori și fructe pe copacii bătrâni. Alte tipuri de răspuns ale plantelor la fertilizarea excesivă includ fasciația sau aplatizarea tulpinilor și necroza internă a scoarței. La răsaduri, efectul nedorit al îngrășământului în exces se manifestă sub formă de creștere apicală excesivă, ceea ce duce la valori scăzute ale raportului dintre părțile subterane și supraterane, drept urmare plantele adesea nu își înrădăcinează bine după transplant.
Utilizarea unor cantități excesive de îngrășăminte este o risipă din punct de vedere economic. De asemenea, este nedorit pentru mediu, deoarece excesul poate fi spălat și ajunge în corpurile de apă sau în apele subterane. De o importanță deosebită este leșierea excesului de azot, de obicei sub formă de nitrat, dar problema poluării mediului poate apărea și atunci când orice element este introdus în cantități excesive.
3.4 Deficit de azot
Cu o lipsă de azot în habitat, creșterea plantelor este inhibată, formarea lăstarilor laterali și slăbirea în cereale este slăbită și se observă frunzele cu frunze mici. În același timp, ramificarea rădăcinilor scade, dar raportul dintre masa rădăcinilor și partea aeriană poate crește. Una dintre manifestările timpurii ale deficitului de azot este culoarea verde pal a frunzelor, cauzată de o slăbire a sintezei clorofilei. Înfometarea prelungită a azotului duce la hidroliza proteinelor și la distrugerea clorofilei, în primul rând în frunzele inferioare, mai bătrâne, și la scurgerea compușilor de azot solubili către frunzele mai tinere și punctele de creștere. Din cauza distrugerii clorofilei, culoarea frunzelor inferioare, în funcție de tipul de plantă, capătă tonuri galbene, portocalii sau roșii, iar cu o deficiență pronunțată de azot pot apărea necroze, uscare și moartea țesuturilor. Înfometarea cu azot duce la o reducere a perioadei de creștere vegetativă și la coacerea mai devreme a semințelor.
3.5 Deficit de fosfor
Un simptom extern al foametei de fosfor este o culoare verde-albăstruie a frunzelor, adesea cu o nuanță violet sau bronz (dovada unei întârzieri a sintezei proteinelor și a acumulării de zaharuri). Frunzele devin mai mici și mai înguste. Creșterea plantelor este suspendată, maturarea culturii este întârziată.
Cu o deficiență de fosfor, rata de absorbție a oxigenului scade, activitatea enzimelor implicate în metabolismul respirator se modifică și unele sisteme de oxidare nemitocondrială (oxidaza acidului glicolic, ascorbat oxidaza) încep să funcționeze mai activ. În condițiile înfometării cu fosfor, procesele de descompunere a compușilor organofosforici și polizaharidelor sunt activate, iar sinteza proteinelor și a nucleotidelor libere este inhibată.
Plantele sunt cele mai sensibile la deficitul de fosfor în stadiile incipiente de creștere și dezvoltare. Nutriția normală cu fosfor într-o perioadă ulterioară accelerează dezvoltarea plantelor (spre deosebire de azot), ceea ce în regiunile sudice reduce probabilitatea ca acestea să cadă sub secetă, iar în nord - sub îngheț.
3.6 Deficit de sulf
Aprovizionarea insuficientă cu sulf a plantelor inhibă sinteza aminoacizilor și proteinelor care conțin sulf, reduce fotosinteza și rata de creștere a plantelor, în special a părților aeriene. În cazurile acute, formarea cloroplastelor este perturbată și este posibilă dezintegrarea acestora. Simptomele deficitului de sulf - albirea și îngălbenirea frunzelor - sunt similare cu cele ale deficitului de azot, dar apar mai întâi pe frunzele cele mai tinere. Acest lucru arată că efluxul de sulf din frunzele mai vechi nu poate compensa furnizarea insuficientă de sulf către plante prin rădăcini.
3.7 Deficit de potasiu
Cu o lipsă de potasiu, îngălbenirea frunzelor începe de jos în sus - de la bătrân la tânăr. Frunzele devin galbene la margini. În viitor, marginile și vârfurile lor devin maro la culoare, uneori cu pete roșii „ruginite”; există o moarte și distrugere a acestor situri. Frunzele par arse. Aprovizionarea cu potasiu este deosebit de importantă pentru organele și țesuturile tinere, în creștere activă. Prin urmare, în timpul înfometării cu potasiu, funcționarea cambiului scade, dezvoltarea țesuturilor vasculare este perturbată, grosimea peretelui celular al epidermei și al cuticulei scade, iar procesele de diviziune și întindere celulară sunt inhibate. Ca urmare a scurtării internodurilor, se pot forma forme de rozetă de plante. Lipsa de potasiu duce la scaderea efectului dominant al mugurilor apicali. Mugurii apicali și apical-laterali încetează să se dezvolte și mor, creșterea lăstarilor laterali este activată și planta ia forma unui tufiș.
Documente similare
Studiul compoziției fizice și chimice a solurilor plantelor de interior, tipuri de îngrășăminte minerale. Semne ale lipsei de minerale în sol. Sfaturi pentru cultivarea plantelor de interior într-un cadru școlar. Boli și dăunători ai plantelor, mijloace de protecție.
lucrare de termen, adăugată 09.03.2014
Rolul mineralelor în activitatea vitală a celulelor și țesuturilor corpului animal. Valoarea macronutrienților pentru organismul animal. Raportul acido-bazic al elementelor din furaj. Utilizarea oligoelementelor în hrănire, ratele de consum zilnic.
rezumat, adăugat 25.10.2009
Clasificarea îngrășămintelor minerale (simple și mixte). Epuizarea solului agricol. Ingrasaminte organice si minerale. Dezvoltarea completă a plantelor atunci când se utilizează îngrășăminte complexe. Influența apei asupra activității vitale a plantelor.
prezentare, adaugat 14.05.2014
Descrierea proteinelor, grăsimilor, carbohidraților, vitaminelor, mineralelor și oligoelementelor. Evaluarea valorii nutritive a furajelor. Metode de studiere a metabolismului în organismul animal, bazate pe legea conservării energiei. Echilibrul de azot, carbon și energie la o vacă.
rezumat, adăugat 15.06.2014
Solurile, condițiile de formare a solului. Caracteristicile îngrășămintelor minerale. Geologie, geomorfologie, clima din vecinătatea râului Sozh. Caracteristici ale solului și condițiilor climatice. Influența îngrășămintelor minerale asupra productivității și compoziției speciilor ierburi.
teză, adăugată 11.03.2012
Dependența calității produselor agricole de conținutul de compuși organici și minerali necesari din acesta. Influența îngrășămintelor minerale (azot, fosfor, potasiu și complex) în diverse combinații asupra dezvoltării și productivității plantelor.
rezumat, adăugat 07.10.2009
Valoarea mineralelor și vitaminelor în corpul porcilor. Utilizarea stimulentelor endogene și a substanțelor biologic active în compoziția premixurilor. Oportunitatea utilizării biostimulanților în dietă (antibiotice, enzime, eleuterococ).
tutorial, adăugat 10/05/2012
Utilizarea îngrășămintelor organice și minerale în districtul Duvansky din Republica Bashkortostan, metode de calcul al dozei de îngrășăminte minerale, planificarea culturilor. Un plan pe termen lung pentru utilizarea îngrășămintelor în rotația culturilor, ținând cont de fertilitatea solului.
lucrare de termen, adăugată 15.07.2009
Semnificația fiziologică a mineralelor în corpul porcilor. Utilizarea picuminului la scroafe în perioada de gestație. Factorii de mediu și influența lor asupra rezistenței naturale și a productivității porcilor. Semăna număr de sânge.
monografie, adăugată 05.10.2012
Dieta unui câine într-un cadru urban. Digestia alimentelor și capacitatea stomacului. Cerințe nutritive și energetice. Rolul grăsimilor în nutriția vitaminelor și metabolismul apei. Simptomele deficitului de acid folic. Funcțiile mineralelor în organism.
Toate substanțele humice se formează ca rezultat al transformării post-mortem (post-mortem) a reziduurilor organice. Transformarea reziduurilor organice în substanțe humice se numește proces de humificare. Are loc în afara organismelor vii, atât cu participarea lor, cât și prin reacții pur chimice de oxidare, reducere, hidroliză, condensare etc.
Spre deosebire de o celulă vie, în care sinteza biopolimerilor se realizează în conformitate cu codul genetic, nu există un program stabilit în procesul de humificare, astfel încât pot apărea orice compuși, atât mai simpli, cât și mai complexi decât biomoleculele originale. Produsele rezultate sunt din nou supuse unor reacții de sinteză sau de descompunere, iar acest proces continuă aproape continuu.
Substanțele humice constituie un grup specific de substanțe de culoare închisă cu molecul mare, formate în timpul descompunerii reziduurilor organice din sol prin sintetizarea țesuturilor vegetale și animale moarte din produse de degradare și degradare. Cantitatea de carbon legată în acizii humici din sol, turbă, cărbuni este de aproape patru ori mai mare decât cantitatea de carbon legat în materia organică a tuturor plantelor și animalelor de pe glob. Dar substanțele humice nu sunt doar produse reziduale ale proceselor de viață, ele sunt produse naturale și cele mai importante ale evoluției comune a substanțelor minerale și a florei Pământului.
Substantele humice pot afecta direct plantele, fiind o sursa de elemente minerale de nutritie (baza de nutrienti). Materia organică a solului conține o cantitate semnificativă de nutrienți, comunitatea vegetală le consumă după ce sunt transformate de microorganismele din sol într-o formă minerală. Sub formă minerală, nutrienții intră în biomasa vegetală.
Substanțele humice pot afecta indirect plantele, adică afectează proprietățile fizico-mecanice, fizico-chimice și biologice ale solului. Oferind un efect complex asupra solului, acestea îmbunătățesc proprietățile fizice, chimice și biologice ale acestuia. Împreună cu aceasta, îndeplinesc o funcție de protecție, legând metalele grele, radionuclizi și toxici organici, împiedicându-le astfel să pătrundă în plante. Astfel, acţionând asupra solului, ele afectează indirect plantele, contribuind la creşterea şi dezvoltarea lor mai activă.
Recent, s-au dezvoltat noi direcții de influență a substanțelor humice asupra plantelor și anume: Plantele sunt heterotrofe care se hrănesc direct cu substanțe humice; Substanțele humice sunt capabile să aibă un efect hormonal asupra plantei, stimulând astfel creșterea și dezvoltarea acesteia.
1. Funcțiile biosferice ale substanțelor humice care afectează dezvoltarea plantelor
În ultimii ani, oamenii de știință au identificat funcțiile biochimice și ecologice generale ale substanțelor humice și influența acestora asupra dezvoltării plantelor. Printre cele mai importante sunt următoarele:
acumulativ- capacitatea substanțelor humice de a acumula rezerve pe termen lung din toate substanțele nutritive, carbohidrați, aminoacizi în diverse medii;
Transport- formarea de compuși organominerale complecși cu metale și oligoelemente care migrează activ în plante;
de reglementare- substanțele humice formează culoarea solului și reglează nutriția minerală, schimbul de cationi, procesele de tamponare și redox din sol;
De protecţie- prin sorbtia de substante toxice si radionuclizi, substantele humice impiedica patrunderea lor in plante.
Combinația tuturor acestor funcții asigură randamente sporite și calitatea necesară a produselor agricole. Este deosebit de important de subliniat efectul pozitiv al acțiunii substanțelor humice în condiții de mediu nefavorabile: temperaturi scăzute și ridicate, lipsă de umiditate, salinitate, acumulare de pesticide și prezența radionuclizilor.
Rolul substanțelor humice ca substanțe active fiziologic este incontestabil. Ele modifică permeabilitatea membranelor celulare, cresc activitatea enzimelor, stimulează procesele de respirație, sinteza proteinelor și carbohidraților. Ele măresc conținutul de clorofilă și productivitatea fotosintezei, care, la rândul său, creează premisele pentru obținerea de produse ecologice.
În utilizarea agricolă a terenului, este necesară o completare constantă a humusului în sol pentru a menține concentrația necesară de substanțe humice.
Până în prezent, această reaprovizionare s-a realizat în principal prin introducerea de compost, gunoi de grajd și turbă. Cu toate acestea, deoarece conținutul de substanțe humice propriu-zise în ele este relativ mic, ratele lor de aplicare sunt foarte mari. Acest lucru crește costurile de transport și alte costuri de producție, care sunt de multe ori mai mari decât costul îngrășământului în sine. În plus, conțin semințe de buruieni, precum și bacterii patogene.
Pentru a obține producții mari și durabile, nu este suficient să ne bazăm pe capacitățile biologice ale culturilor agricole, care, după cum se știe, sunt folosite doar de 10-20%. Desigur, este necesar să se utilizeze soiuri cu randament ridicat, metode eficiente de agro și fitotehnică, îngrășăminte, dar nu se mai poate face fără regulatorii de creștere a plantelor, care până la sfârșitul secolului XX joacă un rol nu mai puțin important decât pesticide și îngrășăminte.
2. Influența nivelului de humus al solului asupra randamentului plantelor agricole
Solurile cu conținut ridicat de humus se caracterizează printr-un conținut mai mare de substanțe active fiziologic. Humusul activează procesele biochimice și fiziologice, crește metabolismul și nivelul general de energie al proceselor din organismul plantei, promovează aportul crescut de nutrienți în acesta, care este însoțit de o creștere a randamentului și de îmbunătățirea calității sale.
În literatură s-a acumulat material experimental, arătând o dependență strânsă a randamentului de nivelul conținutului de humus din sol. Coeficientul de corelație dintre conținutul de humus în sol și randament este de 0,7...0,8 (date de la VNIPTIOU, 1989). Deci, în studiile Institutului de Cercetare a Solului și Agrochimie din Belarus (BelNIIPA), o creștere a cantității de humus din solurile soddy-podzolice cu 1% (în limita schimbării sale de la 1,5 la 2,5 ... 3%) crește randament de boabe de secară de iarnă și orz pentru 10 ... 15 kg / ha. În fermele colective și fermele de stat din regiunea Vladimir, cu un conținut de humus în sol de până la 1%, randamentul de cereale în perioada 1976-1980. nu a depăşit 10 c/ha, la 1,6...2% a fost 15 c/ha, 3,5...4% - 35 c/ha. În regiunea Kirov, o creștere a humusului cu 1% se plătește prin obținerea a încă 3 ... 6 cenți de cereale, în regiunea Voronezh - 2 cenți, în Teritoriul Krasnodar - 3 ... 4 cenți / ha.
Și mai semnificativ este rolul humusului în creșterea rentabilității prin utilizarea cu pricepere a îngrășămintelor chimice, în timp ce eficiența acestuia crește de 1,5 ... 2 ori. Cu toate acestea, trebuie amintit că îngrășămintele chimice aplicate solului provoacă o descompunere crescută a humusului, ceea ce duce la o scădere a conținutului acestuia.
Practica producției agricole moderne arată că creșterea conținutului de humus din sol este unul dintre principalii indicatori ai cultivării acestora. Cu un nivel scăzut de rezerve de humus, aplicarea numai de îngrășăminte minerale nu duce la o creștere stabilă a fertilității solului. Mai mult decât atât, utilizarea unor doze mari de îngrășăminte minerale pe solurile sărace în materie organică este adesea însoțită de efectul lor negativ asupra micro și macroflorei solului, de acumularea de nitrați și alți compuși nocivi în plante și, în multe cazuri, de o scădere a randamentului culturilor. .
3. Efectul substantelor humice asupra plantelor
Acizii humici sunt un produs al transformării biochimice naturale a materiei organice din biosferă. Ele reprezintă partea principală a materiei organice din sol - humus, jucând un rol cheie în ciclul substanțelor din natură și menținând fertilitatea solului.
Acizii humici au o structură moleculară ramificată, incluzând un număr mare de grupuri funcționale și centri activi. Formarea acestor compuși naturali are loc sub influența proceselor fizico-chimice care au loc în sol și a activității organismelor din sol. Sursele de sinteză a acizilor humici sunt reziduurile vegetale și animale, precum și deșeurile microflorei solului.
Astfel, acizii humici sunt acumulatori de materie organică din sol - aminoacizi, glucide, pigmenți, substanțe biologic active și lignină. În plus, componentele anorganice valoroase ale solului sunt concentrate în acizi humici - elemente de nutriție minerale (azot, fosfor, potasiu), precum și oligoelemente (fier, zinc, cupru, mangan, bor, molibden etc.).
Sub influența proceselor naturale care au loc în sol, toate componentele de mai sus sunt incluse într-un singur complex molecular - acizii humici. Varietatea componentelor inițiale pentru sinteza acestui complex determină structura moleculară complexă și, ca urmare, o gamă largă de efecte fizice, chimice și biologice ale acizilor humici asupra solului și plantelor.
Acizii humici, ca parte integrantă a humusului, se găsesc pe aproape toate tipurile de sol. Ele fac parte din combustibilii fosili soli (cărbuni bruni tari și moi), precum și din turbă și sapropel. Cu toate acestea, în stare naturală, acești compuși sunt inactivi și sunt aproape complet într-o formă insolubilă. Fiziologic active sunt doar sărurile formate din acizii humici cu metale alcaline - sodiu, potasiu (humati).
3.1 Influența humaților asupra proprietăților solului
Influența humaților asupra proprietăților fizice ale solurilor
Mecanismul acestui efect variază în funcție de tipul de sol.
Pe solurile argiloase grele, humatele contribuie la respingerea reciprocă a particulelor de argilă prin îndepărtarea sărurilor în exces și distrugerea structurii compacte tridimensionale a argilei. Ca urmare, solul devine mai afânat, excesul de umiditate se evaporă mai ușor, fluxul de aer se îmbunătățește, ceea ce facilitează respirația și avansarea rădăcinilor.
Când sunt aplicate pe soluri ușoare, humatele învăluie și lipesc particulele minerale ale solului, contribuind la crearea unei structuri granulare clocoase rezistente la apă foarte valoroase, care îmbunătățește permeabilitatea la apă și capacitatea de reținere a apei a solului, aerul acestuia. permeabilitate. Aceste caracteristici se datorează capacității acizilor humici de a se gelifica.
Reținerea umidității. Retenția de apă de către humați are loc datorită formării legăturilor de hidrogen între moleculele de apă și grupele încărcate de humați, precum și ionii metalici adsorbiți pe acestea. Ca urmare, evaporarea apei este redusă în medie cu 30%, ceea ce duce la o creștere a absorbției de umiditate de către plante pe soluri aride și nisipoase.
Formarea culorii închise. Humatele colorează solul într-o culoare închisă. Acest lucru este deosebit de important în regiunile reci și temperate, deoarece colorarea închisă îmbunătățește absorbția și stocarea energiei solare de către sol. Ca urmare, temperatura solului crește.
Influența humaților asupra proprietăților chimice ale solurilor și proprietăților umidității solului.
Prin natura lor, acizii humici sunt polielectroliți. În combinație cu particulele de sol organice și minerale, ele formează un complex de absorbție a solului. Dispunând de un număr mare de grupe funcționale diferite, acizii humici sunt capabili să adsorbe și să rețină nutrienții, macro și microelemente care intră în sol. Nutrienții reținuți de acizii humici nu sunt legați de mineralele din sol și nu sunt spălați de apă, fiind într-o stare disponibilă plantelor.
Creșterea capacității tampon a solului. Introducerea de humați crește capacitatea de tampon a solurilor, adică capacitatea solului de a menține un nivel natural de pH chiar și cu un aport în exces de agenți acizi sau alcalini. Deci, atunci când sunt aplicate, humatele sunt capabile să îndepărteze aciditatea excesivă a solurilor, ceea ce în timp face posibilă însămânțarea culturilor sensibile la aciditatea ridicată în aceste câmpuri.
Influența humaților asupra transportului de nutrienți și oligoelemente în plante.
Spre deosebire de acizii humici liberi, humații sunt compuși mobili solubili în apă. Prin adsorbția nutrienților și oligoelementelor, ele facilitează mișcarea lor de la sol la plante.
Odată cu introducerea humaților, există o tendință clară de creștere a conținutului de fosfor mobil (de 1,5-2 ori), potasiu schimbător și azot asimilabil (de 2-2,5 ori) în stratul de sol arabil.
Toate oligoelementele, fiind metale de tranziție (cu excepția borului și iodului), formează complexe chelate mobile cu humați, care pătrund ușor în plante, ceea ce asigură absorbția acestora, în timp ce fierul și manganul, conform oamenilor de știință, sunt absorbite exclusiv sub formă de humați de aceste metale.
Mecanismul presupus al acestui proces este că humații, în anumite condiții, sunt capabili să absoarbă ioni metalici, eliberându-i atunci când condițiile se schimbă. Adăugarea ionilor metalici încărcați pozitiv are loc datorită grupărilor funcționale încărcate negativ ale acizilor humici (carboxilici, hidroxil etc.).
În procesul de absorbție a apei de către rădăcinile plantelor, humatele metalice solubile se apropie de celulele rădăcinii la o distanță apropiată. Sarcina negativă a sistemului radicular depășește sarcina negativă a humaților, ceea ce duce la eliminarea ionilor metalici din moleculele de acid humic și la absorbția ionilor de către membrana celulară.
Mulți cercetători cred că moleculele mici de acizi humici, împreună cu ionii metalici și alți nutrienți atașați acestora, pot fi absorbite și absorbite direct de plantă.
Datorită mecanismelor descrise, nutriția în sol a plantelor se îmbunătățește, ceea ce contribuie la creșterea și dezvoltarea lor mai eficientă.
Influența humaților asupra proprietăților biologice ale solurilor.
Acizii humici sunt surse de fosfați și carbon disponibili pentru microorganisme. Moleculele de acizi humici sunt capabile să formeze agregate mari, pe care există o dezvoltare activă a coloniilor de microorganisme. Astfel, humatele intensifică semnificativ activitatea diferitelor grupe de microorganisme, care sunt strâns legate de mobilizarea nutrienților din sol și de transformarea potențialului fertil în eficient.
Datorită creșterii numărului de bacterii silicate, are loc o completare constantă a potasiului schimbabil absorbit de plante.
Humații cresc numărul de microorganisme din sol care descompun compușii minerali și organici ai fosforului puțin solubili.
Humatele îmbunătățesc aprovizionarea solului cu rezerve de azot asimilabile: numărul bacteriilor amonifiante crește de trei până la cinci ori, în unele cazuri s-a înregistrat o creștere de zece ori a amonifitorilor; numărul bacteriilor nitrificatoare crește de 3-7 ori. Prin îmbunătățirea condițiilor de viață ale bacteriilor care trăiesc liber, capacitatea acestora de a fixa azotul molecular din atmosferă crește de aproape 10 ori.
Ca urmare, solul este îmbogățit cu nutrienți disponibili. În timpul descompunerii materiei organice, se formează o cantitate mare de acizi organici și dioxid de carbon. Sub influența lor, compușii minerali greu accesibile de fosfor, calciu, potasiu, magneziu trec în forme accesibile plantei.
Proprietățile protectoare ale humatelor
Efectul complex al humaților asupra solului oferă proprietățile lor protectoare.
Legarea ireversibilă a metalelor grele și radionuclizilor. Această proprietate a humatelor este deosebit de importantă în condiții de încărcare tehnologică crescută pe sol. Compușii de plumb, mercur, arsen, nichel și cadmiu eliberați în timpul arderii cărbunelui, a funcționării întreprinderilor metalurgice și a centralelor electrice intră în sol din atmosferă sub formă de praf și cenușă, precum și cu gazele de eșapament ale vehiculelor. În același timp, nivelul de poluare cu radiații a crescut semnificativ în multe regiuni.
Când sunt introduși în sol, humații leagă ireversibil metalele grele și radionuclizii. Ca urmare, se formează complexe insolubile, cu mișcare lentă, care sunt îndepărtate din circulația substanțelor în sol. Astfel, humații împiedică pătrunderea acestor compuși în plante și, în consecință, în produsele agricole.
Odată cu aceasta, activarea microflorei de către humați duce la îmbogățirea suplimentară a solului cu acizi humici. Ca urmare, datorită mecanismului descris mai sus, solul devine mai rezistent la poluarea tehnologică.
Accelerarea descompunerii ecotoxicilor organici. Prin activarea activității microorganismelor din sol, humații contribuie la descompunerea accelerată a compușilor organici toxici formați în timpul arderii combustibilului, precum și a pesticidelor.
Compoziția multicomponentă a acizilor humici le permite să absoarbă eficient compușii organici greu accesibile, reducându-le toxicitatea pentru plante și oameni.
3.2 Influența humaților asupra dezvoltării generale a plantelor, semințelor și sistemului radicular
Intensificarea proceselor fizico-chimice și biochimice. Humatele cresc activitatea tuturor celulelor vegetale. Ca urmare, energia celulei crește, proprietățile fizico-chimice ale protoplasmei se îmbunătățesc, metabolismul, fotosinteza și respirația plantelor se intensifică.
Ca urmare, diviziunea celulară este accelerată, ceea ce înseamnă că creșterea generală a plantei se îmbunătățește. Îmbunătățirea nutriției plantelor. Ca urmare a utilizării humatelor, sistemul radicular se dezvoltă activ, crește nutriția rădăcinilor plantelor, precum și absorbția umidității. Intensificarea nutriției rădăcinilor este facilitată de efectul complex al humatelor asupra solului. O creștere a biomasei vegetale și activarea metabolismului duce la o creștere a fotosintezei și la acumularea de carbohidrați de către plante.
Creșterea rezistenței plantelor. Humații sunt activatori nespecifici ai sistemului imunitar. Ca urmare a tratamentului cu humați, rezistența plantelor la diferite boli crește semnificativ. Înmuierea semințelor în soluții de humat este extrem de eficientă pentru a preveni infecțiile semințelor și în special putrezirea rădăcinilor. Odată cu aceasta, tratamentul cu humați crește rezistența plantelor la factorii de mediu negativi - temperaturi extreme, aglomerație, vânturi puternice.
Efectul humatelor asupra semințelor
Datorită tratamentului cu preparate pe bază de substanțe humice, crește rezistența semințelor la boli și leziuni traumatice și are loc o eliberare de infecții de suprafață.
Procesarea semințelor crește germinația, energia germinativă, stimulează creșterea și dezvoltarea răsadurilor.
Astfel, tratamentul crește germinarea semințelor și previne dezvoltarea bolilor fungice, în special a infecțiilor radiculare.
Efectul humatelor asupra sistemului radicular
Permeabilitatea membranei celulelor radiculare crește. Ca urmare, pătrunderea nutrienților și oligoelementelor din soluția de sol în plantă este îmbunătățită. Ca rezultat, nutrienții vin în principal sub formă de complexe cu humați.
Dezvoltarea sistemului radicular se îmbunătățește, crește fixarea plantelor în sol, adică plantele devin mai rezistente la vânturile puternice, spălarea ca urmare a precipitațiilor abundente și proceselor de eroziune.
Mai ales eficient asupra culturilor cu un sistem radicular subdezvoltat: grâu de primăvară, orz, ovăz, orez, hrișcă.
Dezvoltarea sistemului radicular intensifică absorbția umidității și oxigenului de către plantă, precum și nutriția solului.
Ca urmare, sinteza aminoacizilor, zaharurilor, vitaminelor și acizilor organici este îmbunătățită în sistemul radicular. Schimbul de substante intre radacini si sol este intensificat. Acizii organici secretati de radacini (carbonici, malici etc.) afecteaza in mod activ solul, crescand disponibilitatea nutrientilor si microelementelor.
4. Concluzie
Substanțele humice, fără îndoială, au un impact asupra creșterii și dezvoltării plantelor. Materia organică din sol servește ca sursă de nutrienți pentru plante. Microorganismele, substanțele humice în descompunere, furnizează plantelor substanțe nutritive sub formă minerală.
Substanțele humice au un impact semnificativ asupra complexului de proprietăți ale solului, afectând astfel indirect dezvoltarea plantelor.
Substanțele humice, îmbunătățind proprietățile fizico-chimice, chimice și biologice ale solului, stimulează creșterea și dezvoltarea mai intensă a plantelor.
De mare importanță, în prezent, datorită creșterii intense a impactului antropic asupra mediului înconjurător în general, și asupra solului în special, este și funcția protectoare a substanțelor humice. Substanțele humice leagă substanțele toxice și radionuclizii și, ca urmare, contribuie la producerea de produse ecologice.
Substanțele humice au cu siguranță un efect benefic atât asupra solului, cât și asupra plantelor.
Lista literaturii folosite.
- Aleksandrova L.N. Materia organică din sol și procesele de transformare a acesteia. L., Știință, 1980,
- Orlov D.S. Acizii humici ai solurilor și teoria generală a humificării. M.: Editura Universității de Stat din Moscova, 1990.
- Ponomareva V.V., Plotnikova T.A. Formarea humusului și a solului. L., Știință, 1980,
- Tyurin I.V. Materia organică din sol și rolul acesteia în formarea și fertilitatea solului. Doctrina humusului din sol. Selhozgiz, 1967.
- Tate R., III. Materia organică din sol. M.: Mir, 1991..
- Khristeva L.A. Efectul de stimulare al acidului humic asupra creșterii plantelor superioare și a naturii acestui fenomen. 1957.
- Substanțe humice din biosferă. Ed. D.S. Orlov. Moscova: Nauka, 1993.
Influența substanțelor chimice asupra creșterii și dezvoltării plantelor. Completat de: Ignatieva Victoria, elevă în clasa a VI-a Conducător: Putina Yu.K., profesor de biologie și chimie Instituția municipală de învățământ de stat „Școala secundară Nizhnesanarskaya a districtului municipal Troitsky din regiunea Chelyabinsk 2017
Scop: studierea efectului substanțelor chimice asupra creșterii și dezvoltării plantelor. Obiective: Studierea literaturii disponibile pe această temă; Pentru a se familiariza cu metodele disponibile pentru studiul efectului substanțelor chimice asupra creșterii și dezvoltării plantelor. Faceți o concluzie despre efectele substanțelor chimice pe baza propriilor cercetări. Elaborați recomandări pentru îmbunătățirea condițiilor de cultivare a culturilor. Ipoteza: Presupunem ca substantele chimice vor afecta negativ cresterea si dezvoltarea plantelor.
Obiectul de studiu: Ceapa bulbuca, Fasole comuna Subiect de studiu: efectul substantelor chimice asupra plantelor.
Procedura de eșantionare chimică
Pentru studiul efectului substanțelor chimice au fost prelevate 6 probe: Nr. 1 - sulfat de cupru CuSO4 * 5H2O Nr. 2 - sulfat de zinc ZnSO4 * 7H2O Nr. 3 - sulfat de fier FeSO4 * 7H2O Nr. 4 - permanganat de potasiu KMnO4 Nr. 5 - sulfat de plumb PbSO4 nr. 6 - proba de control (fără substanțe chimice adăugate)
Rezultatele studiului probelor martor Eșantionul de control nr. 6 (bulbul de ceapă) dezvoltarea se desfășoară intens cu formarea multor rădăcini adventive) Proba de control nr. 6 (plantă de fasole) - creșterea și dezvoltarea sunt în limite normale
Rezultatele studiului probelor de testare sub influența sulfatului de cupru Proba nr. 1 Apariția unui număr mic de rădăcini, creșterea lor se oprește curând, se întunecă. Proba nr. 1 După adăugarea unei soluții de sulfat de cupru, frunzele s-au ondulat imediat, planta a murit până la sfârșitul primei săptămâni a experimentului
Rezultatele studiului probelor de testare sub influența sulfatului de zinc Proba nr. 2 Apariția unui număr mare de rădăcini, creșterea lor este nesemnificativă. Proba nr. 2 În plantă, după adăugarea unei soluții de sulfat de zinc, frunzele s-au dezvoltat de obicei în prima săptămână de experimente, apoi, odată cu creșterea concentrației soluției, frunzele s-au îngălbenit, s-au ondulat.
Rezultatele studiului probelor de testare sub influența sulfatului de fier Proba nr. 3 Apariția unui număr mic de rădăcini, creșterea lor se oprește curând, se întunecă. Proba nr. 3. planta a dezvoltat trei frunze, dar apoi au început să se îndoaie și să se îngălbenească
Rezultatele studiului probelor de testare atunci când sunt expuse la permanganat de potasiu Proba nr. 4 Bulbul cu adaos de soluție de permanganat de potasiu (nr. 4) s-a dezvoltat slab, rădăcinile 1-2 mm, apoi creșterea s-a oprit Proba nr. 4 Planta a pierdut 3 frunze in ziua 4, apoi restul s-au ofilit
Rezultatele studiului probelor de testat sub influența sulfatului de plumb Proba nr. 5 Bulbul avea un număr suficient de rădăcini, dar de dimensiuni reduse. Planta de fasole avea frunze mari, dar de o culoare palidă, care și ele s-au ondulat ușor la sfârșitul a 2 săptămâni
Proba de control (nr. 6) a avut chiar celule ușoare fără semne de deformare.
Celulele de ceapă din proba experimentală cu adaos de sulfat de fier (nr. 3) au avut o structură uniformă, dar citoplasma lor era de culoare închisă.
Celulele de ceapă dintr-o probă experimentală cu adăugare de permanganat de potasiu (nr. 4) au devenit albastre. Celulele aveau o structură uniformă.
Concluzii: Un exces de sulfat de fier colorează celulele într-o culoare închisă și încetinește creșterea sistemului radicular. Permanganatul de potasiu are același efect. Excesul de sulfat de cupru distruge celulele plantelor și oprește creșterea acesteia.
LUCRARE DE CURS
Influența diferitelor tipuri de tratare a semințelor asupra creșterii și dezvoltării plantelor
Introducere
Problema tratarii semințelor înainte de însămânțare, în ciuda numeroaselor studii, rămâne relevantă și deschisă până acum. Interesul este generat de perspectiva utilizării diferitelor tipuri de tratare a semințelor în agricultură pentru a crește productivitatea plantelor și a obține randamente mai mari.
În timpul depozitării, semințele îmbătrânesc, calitatea și germinarea semințelor scad, prin urmare, într-un lot de semințe depozitate câțiva ani, există semințe puternice, slabe (vii, dar nu germinate) și semințe moarte. Metode cunoscute de tratare a semințelor înainte de însămânțare, care pot crește germinarea semințelor pierdute în timpul depozitării. Radiațiile ionizante în doze mici, sonorizarea, tratamentele termice și cu unde de șoc pe termen scurt, expunerea la câmpuri electrice și magnetice, iradierea cu laser, înmuierea prealabilă în soluții de substanțe biologic active și altele pot crește germinarea semințelor și randamentul cu 15-25% .
După cum știți, îngrășămintele minerale sunt folosite pentru a crește productivitatea, sunt aplicate convenabil pe sol, acest proces este mecanizat. Utilizarea îngrășămintelor minerale determină creșterea accelerată a plantelor și creșterea randamentelor. Cu toate acestea, nitrații și nitriții, care nu sunt periculoși pentru plante, dar periculoși pentru oameni, se formează adesea în paralel. În plus, există consecințe mai grave ale utilizării îngrășămintelor minerale asociate cu modificările structurii solului. Ca urmare, îngrășămintele sunt spălate din straturile superioare ale solului către cele inferioare, unde componentele minerale nu mai sunt disponibile plantelor. Apoi, îngrășămintele minerale intră în apele subterane și sunt transportate în corpurile de apă de suprafață, poluând semnificativ mediul. Utilizarea îngrășămintelor organice este mai ecologică, dar în mod clar nu sunt suficiente pentru a satisface nevoia umană de a crește productivitatea.
Metodele fizice sigure din punct de vedere ecologic de biostimulare a semințelor sunt foarte promițătoare. În prezent, s-a dovedit experimental că obiectele biologice sunt capabile să răspundă sensibil la impactul câmpurilor electromagnetice externe. Această reacție poate avea loc la diferite niveluri structurale ale unui organism viu - de la moleculară și celulară până la organismul în ansamblu. Sub influența undelor electromagnetice din gama milimetrică în celulele obiectelor biologice, sunt activate procesele de biosinteză și diviziune celulară, sunt restabilite conexiunile și funcțiile perturbate din cauza bolilor, sunt sintetizate suplimentar substanțele care afectează starea imunitară a organismului.
Până în prezent, au fost dezvoltate un număr mare de diverse instalații de iradiere și metode de activare a semințelor. Cu toate acestea, nu au primit o distribuție largă, deși sunt mai avansate din punct de vedere tehnologic, sigure pentru mediu și mult mai ieftine în comparație cu metodele chimice. Unul dintre motivele acestei situații este acela că metodele existente de tratare a semințelor cu radiații nu dau rezultate constant ridicate. Acest lucru se datorează faptului că în metodele existente de tratare înainte de însămânțare, caracteristicile calitative și cantitative ale radiațiilor nu sunt optimizate.
Scopul studiului - să studieze influența diferitelor tipuri de tratament al semințelor înainte de însămânțare asupra creșterii și dezvoltării plantelor.
În acest sens, următoarele sarcini :
Să studieze influența substanțelor chimice asupra creșterii și dezvoltării plantelor;
· să studieze influența tratamentului electromagnetic (biofizic) asupra proceselor de creștere la plante;
· dezvăluie efectul iradierii cu laser asupra germinării semințelor de orz.
1. Tratarea semințelor înainte de însămânțare și efectul acestuia asupra creșterii și dezvoltării plantelor
1.1 Efectul substanțelor chimice asupra creșterii și dezvoltării plantelor
iradierea cu laser a semințelor de orz
Cea mai importantă și eficientă parte a tratamentului este pansamentul chimic sau pentru semințe.
Chiar și acum 4 mii de ani, în Egiptul și Grecia antic, semințele erau înmuiate în suc de ceapă sau mutate în timpul depozitării cu ace de chiparos.
În Evul Mediu, odată cu dezvoltarea alchimiei și, datorită acesteia, chimiștii au început să înmuie semințele în sare de rocă și potasiu, vitriol albastru și săruri de arsenic. În Germania, cele mai simple metode erau populare - păstrarea semințelor în apă fierbinte sau într-o soluție de gunoi de grajd.
La începutul secolului al XVI-lea, s-a observat că semințele care au fost în apa mării în timpul unui naufragiu produceau culturi care au fost mai puțin afectate de pulverul dur. Mult mai târziu, acum 300 de ani, eficacitatea tratamentului chimic al semințelor înainte de însămânțare a fost dovedită științific în cursul experimentelor omului de știință francez Thiele, care a investigat efectul tratamentului semințelor cu sare și var asupra răspândirii prin semințe de durere. negru de fum.
La începutul secolului al XIX-lea, utilizarea preparatelor cu arsenic ca fiind periculoase pentru viața umană a fost interzisă, dar la începutul secolului al XX-lea au început să folosească substanțe care conțin mercur, care au fost interzise abia în 1982 și abia în 1982. în Europa de Vest.
Abia în anii 1960 au fost dezvoltate fungicide sistemice pentru pretratarea semințelor, iar țările industrializate au început să le folosească în mod activ. Începând cu anii 90, au fost utilizate complexe de insecticide și fungicide moderne extrem de eficiente și relativ sigure.
În funcție de tehnologia de tratare a semințelor, se disting trei tipuri de tratare a semințelor: pansare simplă, drajare și încrustare.
Pansamentul standard este cel mai comun și tradițional mod de tratare a semințelor. Cel mai adesea folosit în grădini și ferme, precum și în producția de semințe. Crește greutatea semințelor cu cel mult 2%. Dacă compoziția filmogenă acoperă complet semințele, greutatea acestora poate crește cu până la 20%.
Incrustare - semintele sunt acoperite cu substante lipicioase care asigura fixarea substantelor chimice pe suprafata lor. Semințele tratate pot deveni de 5 ori mai grele, dar forma nu se schimbă.
Acoperire - substanțele acoperă semințele cu un strat gros, crescându-le greutatea de până la 25 de ori și schimbând forma în sferică sau eliptică. Cea mai „puternică” drajare (peletizare) face semințele de până la 100 de ori mai grele.
Pentru tratarea semințelor de cereale se folosesc cel mai activ preparatele Raxil, Premix, Vincite, Divident, Colfugo Super Color. Acestea sunt fungicide sistemice care ucid sporii de piatră, pulberul praf și dur, nematozi care luptă eficient cu Fusarium, Septoria și putregaiul rădăcinilor. Acestea sunt produse sub formă de lichide, pulberi sau suspensii concentrate și sunt utilizate pentru tratarea semințelor în aparate speciale la doza de 0,5-2 kg la 1 tonă de semințe.
În gospodăriile private și agricole, utilizarea substanțelor chimice puternice nu este întotdeauna justificată. Cantități relativ mici de semințe mici ale culturilor legumicole sau ornamentale, precum gălbenele, morcovii sau roșiile, pot fi tratate cu substanțe mai puțin toxice. Este important nu numai și nu atât de mult să distrugi inițial întreaga infecție pe semințe, ci să formezi rezistență la boli în plantă în stadiul embrionului de semințe, adică o imunitate puternică.
La începutul germinării sunt benefice și stimulentele de creștere, care vor încuraja dezvoltarea unui număr mare de rădăcini laterale la plante, creând un sistem radicular puternic. Stimulenții de creștere a plantelor, care intră în embrion înainte de germinare, determină transportul activ al nutrienților către părțile aeriene ale plantei. Semințele tratate cu astfel de preparate germinează mai repede, germinarea lor crește. Răsadurile devin mai rezistente nu numai la boli, ci și la temperaturi extreme, lipsă de umiditate și alte condiții stresante. Consecințele mai îndepărtate ale pre-tratării adecvate cu preparate înainte de însămânțare sunt considerate a fi o creștere a randamentului și o reducere a timpului de coacere.
Multe preparate pentru tratarea semințelor înainte de însămânțare sunt create pe bază humică. Sunt o soluție apoasă concentrată (până la 75%) de acizi humici și humați, potasiu și sodiu, saturată cu un complex de minerale necesare plantei, care poate fi folosit și ca îngrășământ. Astfel de preparate sunt produse pe baza de turba, fiind extractul ei de apa.
Z.F. Rakhmankulova și colab. au studiat efectul pre-semănării semințelor de grâu (Triticum aestivum L.) cu acid salicilic (SA) de 0,05 mm asupra conținutului său endogen și a raportului dintre formele libere și legate în lăstarii și rădăcinile răsadurilor. Pe parcursul a două săptămâni de creștere a răsadurilor, s-a observat o scădere treptată a conținutului total de SA în lăstari; nu s-au găsit modificări la rădăcini. În același timp, a existat o redistribuire a formelor SA în lăstari - o creștere a nivelului formei conjugate și o scădere a formei libere. Tratamentul înainte de însămânțare a semințelor cu salicilat a dus la scăderea conținutului total de SA endogen atât în lăstari, cât și în rădăcinile răsadurilor. Conținutul de SA liber a scăzut cel mai intens în lăstari și ceva mai puțin în rădăcini. S-a presupus că o astfel de scădere a fost cauzată de o încălcare a biosintezei SA. Aceasta a fost însoțită de o creștere a masei și lungimii lăstarilor și în special a rădăcinilor, stimularea respirației totale întunecate și o modificare a raportului căilor respiratorii. S-a observat o creștere a proporției căii de respirație a citocromului la rădăcini, iar la lăstari s-a observat o creștere a ponderii căii alternative rezistente la cianuri. Sunt prezentate modificări ale sistemului antioxidant al plantelor. Gradul de peroxidare a lipidelor a fost mai pronunțat la lăstari. Sub influența pretratării cu SA, conținutul de MDA în lăstari a crescut de 2,5 ori, în timp ce în rădăcini a scăzut de 1,7 ori. Din datele prezentate rezultă că natura și intensitatea efectului SA exogen asupra creșterii, echilibrului energetic și statusului antioxidant al plantelor pot fi asociate cu modificări ale conținutului său în celule și cu redistribuirea între formele SA libere și conjugate.
E.K. Eskov, în experimentele de producție, a studiat efectul pre-semănării semințelor de porumb cu nanoparticule de fier asupra intensificării creșterii și dezvoltării, crescând randamentul de masă verde și boabe a acestei culturi. Ca urmare, a avut loc o intensificare a proceselor fotosintetice. Conținutul de Fe, Cu, Mn, Cd și Pb în ontogeneza porumbului a variat foarte mult, dar adsorbția nanoparticulelor de Fe în stadiile inițiale de dezvoltare a plantelor a afectat scăderea conținutului acestor elemente chimice în boabele de coacere, care a fost însoțită. printr-o modificare a proprietăților sale biochimice.
Astfel, tratarea înainte de însămânțare a semințelor cu substanțe chimice este asociată cu costuri ridicate cu forța de muncă și cu o capacitate scăzută de fabricație a procesului. În plus, utilizarea pesticidelor în scopul dezinfectării semințelor dăunează mult mediului.
1.2 Efectul tratamentului electromagnetic (biofizic) asupra proceselor de creștere la plante
În contextul unei creșteri puternice a costului purtătorilor de energie, al poluării tehnologice a agroecosistemelor, este necesar să se caute resurse materiale și energetice ecologice și rentabile ca alternativă la mijloacele costisitoare și nesigure din punct de vedere ecologic de creștere a productivității, îmbunătățind în același timp calitatea culturilor.
Metodele și metodele tehnologice existente de stimulare înainte de însămânțare a semințelor, bazate pe utilizarea de substanțe chimice foarte toxice, sunt asociate cu costuri ridicate ale forței de muncă și cu o fabricabilitate scăzută a procesului de tratare a semințelor. În plus, utilizarea pesticidelor în scopul dezinfectării semințelor dăunează mult mediului. Când semințele tratate cu fungicide sunt introduse în sol, pesticidele sub influența vântului și a ploii sunt transportate în corpurile de apă, răspândite pe suprafețe vaste, ceea ce poluează mediul și dăunează naturii.
De cel mai mare interes pentru obținerea de produse ecologice sunt factorii fizici ai câmpului electromagnetic, cum ar fi radiațiile gamma, razele X, ultravioletele, vizibile optice, infraroșii, radiațiile cu microunde, frecvența radio, câmpurile magnetice și electrice, expunerea la particule alfa și beta. , ionii diferitelor elemente , influența gravitațională etc. Utilizarea iradierii cu raze gamma și X este periculoasă pentru viața umană și, prin urmare, nepotrivită pentru utilizare în agricultură. Utilizarea iradierii cu ultraviolete, microunde și radiofrecvență provoacă probleme în timpul funcționării. Relevant este studiul impactului câmpurilor electromagnetice în cultivarea cerealelor, a solaanei, a semințelor oleaginoase, a leguminoaselor, a pepenilor și a rădăcinilor.
Acțiunea câmpurilor magnetice este asociată cu efectul acestora asupra membranelor celulare. Impactul dipolului stimulează aceste modificări ale membranelor, îmbunătățește activitatea enzimelor. În plus, s-a stabilit de alți autori că, în urma unui astfel de tratament, în semințe au loc o serie de procese, ducând la creșterea permeabilității învelișului semințelor, iar fluxul de apă și oxigen în semințe este accelerat. Ca urmare, activitatea enzimatică este îmbunătățită, în primul rând, enzimele hidrolitice și redox. Acest lucru asigură o aprovizionare mai rapidă și mai completă cu nutrienți a embrionului, accelerarea ratei de diviziune celulară și activarea proceselor de creștere în general. La plantele crescute din semințe tratate, sistemul radicular se dezvoltă mai intens și trecerea la fotosinteză este accelerată, adică. se creează o bază solidă pentru creșterea și dezvoltarea în continuare a plantelor.
Toate acestea contribuie la procesul vegetativ, accelerează creșterea acestuia.
Au fost realizate noi nanotehnologii de tratare a semințelor înainte de însămânțare cu microunde și de combatere a dăunătorilor ca alternativă la metodele chimice. Pentru dezinsecția cerealelor și semințelor s-a folosit un mod de tratament cu microunde pulsat care, datorită intensității ultra-înalte a EMF în puls, asigură moartea insectelor dăunătoare. S-a stabilit că pentru un efect 100% de dezinsecție cu microunde este necesară o doză de cel mult 75 MJ la 1 tonă de semințe. Dar astăzi aceste tehnologii nu pot fi utilizate direct în complexul agroindustrial, deoarece doar dezvoltarea lor este în curs de dezvoltare, iar costul estimat al introducerii lor în producție este foarte mare. Printre practicile agricole promițătoare, care au un efect stimulativ asupra creșterii și dezvoltării plantelor, ar trebui să se includă utilizarea câmpurilor electrice și magnetice, care sunt utilizate atât la pregătirea înainte de însămânțare a semințelor, cât și în timpul sezonului de vegetație a plantelor prin creșterea rezistenta plantelor la factorii de stres, crescand factorul de utilizare a substantelor nutritive din sol, ceea ce duce la cresterea randamentelor culturilor. Sa dovedit influența pozitivă a câmpului electromagnetic asupra calităților de semănat și de producție a semințelor culturilor de cereale.
Tratarea electromagnetică a semințelor, în comparație cu o serie de alte metode de tratament, nu este asociată cu operațiuni intensive și costisitoare de muncă, nu are un efect dăunător asupra personalului de întreținere (cum ar fi tratamentul chimic sau cu radionuclizi) sau utilizarea pesticidelor, nu nu se administrează doze care sunt letale pentru semințe în timpul procesării, este un proces foarte tehnologic și ușor de automatizat, impactul este ușor și precis dozat, este un tip de prelucrare ecologic, se încadrează ușor în practicile agricole utilizate în prezent. De asemenea, este important ca plantele crescute din semințe tratate să nu aibă alte modificări patologice și mutații induse. Se demonstrează că impactul unui câmp electromagnetic mărește numărul de tulpini productive, numărul de spiculete, lungimea medie a plantelor și a spițelor, crește numărul de boabe în spion și, în consecință, masa boabelor. Toate acestea duc la o creștere a randamentului cu 10-15%.
G.V. Novitskaya a studiat efectul unui câmp magnetic orizontal (CMF) constant slab cu o putere de 403 A/m asupra compoziției și conținutului lipidelor polare și neutre și a AG constituenți ai acestora în frunzele principalelor tipuri de orientare magnetică (MOT) ale ridiche (Raphanus sativus L., var. radicula D. C.) soiuri Roz-rosu cu varf alb: nord-sud (NS) si vest-est (WE), in care sunt situate planurile de orientare ale brazdelor radiculare. de-a lungul și, respectiv, peste meridianul magnetic. Sub acțiunea PMF primăvara, conținutul total de lipide din frunzele MOT NS a scăzut, în timp ce în frunzele MOT WE a crescut; toamna, dimpotriva, a crescut continutul total de lipide din frunzele SL MOT, in timp ce cel al WE MOT a scazut. Primăvara, raportul dintre fosfolipide și steroli, indicând indirect o creștere a fluidității dublului strat lipidic al membranelor, a crescut la plantele ambelor MOT, în timp ce toamna a crescut doar în MOT-urile CL. Conținutul relativ de acizi grași nesaturați, inclusiv acizii linolenic și linoleic, la martor a fost mai mare în SR MOT comparativ cu NC MOT. Sub acțiunea PMP, conținutul acestor acizi în lipidele frunzelor SL MOT a crescut, în timp ce cel al WE MOT a rămas neschimbat. Astfel, PMF orizontal slab a avut un efect diferit, uneori opus, asupra conținutului de lipide al frunzelor de SN și WE MOT de ridiche, care, aparent, este cauzat de sensibilitatea lor diferită la acțiunea câmpului, asociată cu particularitățile starea lor fiziologică.
În plus, G.V. Novitskaya și colab. au studiat efectul PMF cu o putere de 403 A/m asupra compoziției și conținutului de lipide polare (cap) și neutre și acizii grași constituenți ai acestora izolați din 3, 4 și 5 frunze de plante de ceapă (Allium sera L .) cv. folosind metodele TLC și GLC. Plantele crescute în câmpul magnetic natural al Pământului au servit drept control. Sub acțiunea PMF, cele mai mari modificări ale conținutului de lipide s-au întâlnit la a patra frunză a cepei: conținutul total de lipide a crescut, în special, lipidele polare (glico- și fosfolipide), în timp ce cantitatea de lipide neutre a scăzut sau ramas neschimbat. Raportul fosfolipide/steroli a crescut, indicând o creștere a fluidității stratului dublu lipidic al membranelor. Sub influența PMP, proporția de acid linolenic a crescut și a crescut și conținutul relativ al acizilor grași nesaturați totali. Efectul PMP asupra compoziției și conținutului de lipide al celei de-a treia și a cincea frunze de ceapă a fost mai puțin pronunțat, ceea ce indică o sensibilitate diferită a frunzelor de ceapă de diferite vârste la acțiunea câmpului. Se concluzionează că modificările PMF slabe din trecutul modificări evolutive-istorice ale intensității câmpului magnetic al Pământului pot afecta compoziția biochimică și procesele fiziologice ale plantelor.
În cursul studiilor privind influența unui câmp magnetic alternant (AMF) cu o frecvență de 50 Hz asupra dinamicii desfășurării frunzelor de cotiledon, compoziția și conținutul de lipide polare și neutre și acizii grași constituenți ai acestora în 5 zile -rasadurile vechi de ridichi crescute la lumina si la intuneric ( Raphanus sativus L. var. radicula DL) soiul trandafir-rosu cu varful alb, s-a constatat ca PMF a slabit efectul inhibitor al luminii asupra dinamicii desfasurarii frunzelor de cotiledon. La lumină în PMP, conținutul total de lipide, conținutul de lipide polare și neutre din răsaduri a fost mai mare decât la martor. Dintre lipidele polare, conținutul total de glico- și fosfolipide a crescut; dintre lipidele neutre, conținutul de triacilgliceroli a crescut. Raportul dintre fosfolipide și steroli (PL/ST) a crescut. Pe întuneric, în PMF, conținutul total de lipide, precum și de lipide neutre în răsaduri, a fost mai mic decât la martor, iar raportul PL/ST a scăzut. La martor nu s-au constatat diferențe în ceea ce privește conținutul total relativ de acizi grași nesaturați la lumină și la întuneric; conținutul de acid linolenic din răsaduri a fost mai mare la lumină decât la întuneric. Sub acțiunea PMF, conținutul de acid linolenic în lumină a scăzut, a crescut la întuneric, iar acidul erucic a scăzut la lumină. Raportul dintre acizii grași nesaturați și saturați a scăzut atât la lumină, cât și la întuneric. Se ajunge la concluzia că PMF cu o frecvență de 50 Hz a modificat semnificativ conținutul de lipide din răsadurile de ridiche la lumină și pe întuneric, acționând ca un factor corector.
Astfel, studiile multor autori au stabilit că sub influența unui câmp electromagnetic se mobilizează forțele și se eliberează rezervele de energie ale organismului, se activează procesele fiziologice și biochimice în stadiile incipiente ale germinării semințelor, se înregistrează o creștere a procesele intra-metabolice și o creștere constantă a energiei germinative, germinația, puterea, creșterea inițială, supraviețuirea primăvara-vară, care afectează favorabil întreaga perioadă ulterioară de dezvoltare a plantei.
Cu toate acestea, nu au primit o distribuție largă, deși sunt mai avansate din punct de vedere tehnologic, sigure pentru mediu și mult mai ieftine în comparație cu metodele chimice. Unul dintre motivele acestei situații este acela că metodele existente de tratare a semințelor cu radiații nu dau rezultate constant ridicate. Acest lucru se datorează modificărilor condițiilor externe, eterogenității materialului de semințe și cunoașterii insuficiente a esenței interacțiunii celulelor de semințe cu câmpurile electromagnetice și sarcinile electrice.
1.3 Efectul iradierii cu laser asupra creșterii și dezvoltării plantelor
Din cele mai vechi timpuri, îmbunătățirea fertilității solului a fost considerată pe bună dreptate cea mai importantă condiție pentru creșterea productivității producției vegetale. Sume uriașe de bani și eforturi ale oamenilor de știință din întreaga lume sunt cheltuite pentru reabilitarea terenurilor, irigarea și chimizarea agriculturii. Cu toate acestea, tristul paradox al progresului în chimiizarea agriculturii este că, după utilizarea excesivă a nitraților, fosfaților, pesticidelor, regulatorilor sintetici de creștere, o umbră malefică urmează otrăvirea culturilor, alimentelor, apei, o amenințare pentru sănătatea și viața umană. În consecință, are loc o intensificare a dezvoltării de noi modalități și metode de intensificare a productivității producției vegetale.
Sub forma uneia dintre aceste metode, este prezentată o radiație laser sau laser. Deoarece centrele științifice moderne au început să acorde o mare atenție tehnologiilor moderne de cultivare a culturilor, în astfel de condiții au fost dezvoltate o serie de metode de influențare a culturilor agricole cu diverși factori fizici care au un efect stimulativ asupra creșterii și dezvoltării plantelor și, în cele din urmă, , pe randamentul culturilor în sine. Plantele sau semințele lor au început să fie plasate în câmpuri magnetice sau electrice puternice, pentru a influența culturile cu radiații ionizante sau plasmă, precum și iradierea luminii concentrate a soarelui - lumina surselor moderne de radiații create artificial - laserele.
Acțiunea prelucrării cu laser în ansamblu poate fi numită specifică, deoarece este un factor pozitiv din punct de vedere al ecologiei și siguranței pentru mediu, deoarece nu sunt introduse elemente străine în natură atunci când sunt expuse la aceasta.
Metoda de expunere la un laser concentrează un număr suficient de avantaje în comparație cu alte metode fizice și chimice existente de pregătire a semințelor înainte de însămânțare și anume:
1) o creștere stabilă a randamentelor culturilor pe fondul diferitelor condiții pedoclimatice;
2) îmbunătățirea calității produselor agricole (creșterea conținutului de zaharuri, vitamine, proteine și gluten);
3) posibilitatea reducerii ratei de însămânțare cu 10-30% prin creșterea germinării în câmp a semințelor și intensificarea proceselor de creștere (în funcție de soi, tip de cultură, frecvență de prelucrare);
4) creșterea rezistenței plantelor la deteriorarea prin diferite boli;
5) inofensivitatea procesării pentru semințe și personalul de service.
Totuși, efectul pozitiv al iradierii cu laser a semințelor și plantelor are și o parte de dezavantaje care trebuie luate în considerare. Astfel, amploarea efectului de activare și reproductibilitatea acestuia depind de starea semințelor, care este influențată de mulți factori naturali și necontrolați în timpul depozitării și iradierii. În plus, în anumite condiții, iradierea semințelor cu doze optime poate să nu afecteze deloc activitatea plantelor și chiar să aibă un efect deprimant.
F.D. Samuilov a studiat microvâscozitatea mediului apos în embrionii și endospermul semințelor de porumb (Zea mays L.) iradiate folosind un laser electronic Lvov-1 folosind o sondă de spin. Conform parametrilor spectrelor EPR ai radicalilor nitroxil (sonde) absorbiți de semințe cu apă în timpul umflăturii, s-au determinat timpii de corelație ai difuziei rotative a sondei C în embrioni și endospermul semințelor. S-a constatat o scădere a C a sondelor din embrionii semințelor iradiate în comparație cu semințele neiradiate și s-a stabilit dependența valorii C de timpul de umflare a semințelor. Se ajunge la concluzia că în celulele embrionilor de semințe sub acțiunea iradierii laser, microvâscozitatea mediului apos scade și mobilitatea sondelor crește. Efectul iradierii asupra sondelor C din endospermul semințelor se manifestă într-o măsură mai mică și este, de asemenea, însoțit de o creștere a mobilității sondei.
Astfel, metoda de tratare cu laser are o serie de avantaje față de metodele fizice și chimice de pregătire a semințelor înainte de însămânțare. Acestea includ: îmbunătățirea calității produselor agricole (creșterea conținutului de zaharuri, vitamine, proteine și gluten); posibilitatea reducerii ratei de însămânțare cu 10-30% prin creșterea germinării în câmp a semințelor și intensificarea proceselor de creștere; inofensivitatea procesării pentru semințe și personalul de service; durată scurtă de expunere. Dar tratarea semințelor cu laser este foarte costisitoare și, prin urmare, nu este utilizată pe scară largă în fermă. Iradierea gamma face posibilă accelerarea germinării semințelor unor plante cultivate, crește germinația câmpului și numărul de tulpini productive și, ca urmare, randamentele (până la 13%). Dezavantajele includ dependența eficacității iradierii preînsămânțate de condițiile meteorologice în timpul sezonului de vegetație, un impact negativ asupra unui număr de trăsături economice ale plantelor și o scădere a intensității regimului respirator al plantelor. Principalul dezavantaj al acestei metode de stimulare este că o creștere a dozei de tratament poate fi fatală.
2. Obiecte şi metode de cercetare
Cercetarea a fost efectuată la Departamentul de Botanică și Fundamente ale Agriculturii, Universitatea Pedagogică de Stat din Belarus. M. Tanka și Facultatea de Fizică a BSU.
2.1 Obiectul de studiu
Obiectul studiului îl reprezintă semințele soiului de orz Yakub. Această varietate de selecție din Belarus, obținută de Întreprinderea Unitară Republicană „Centrul Științific și Practic al Academiei Naționale de Științe din Belarus pentru Agricultură” și inclusă în Registrul de Stat în 2002.
Caracteristici morfologicesoiuri. O plantă în faza de măcinare de tip intermediar. Tulpina are o înălțime de până la 100 cm. Poziția urechii este semi-erectă. Spiculetul este pe două rânduri, cilindric, cu lungimea de până la 10 cm, cu 26-28 de spiculețe pe spișă. Arzuri de lungime medie în raport cu urechea. Granule peliculoase. Şanţul ventral nu este pubescent. Stratul de aleuronă al cariopsei este ușor colorat. Tip de dezvoltare - primăvară.
Caracteristici economice și biologicesoiuri. Soi de cereale. Dimensiunea boabelor - mare (greutate de 1000 de boabe - 45-50 g). Soi bogat în proteine (conținut mediu de proteine 15,4%, randament proteic la hectar până la 6,0 q). Soi mediu târziu. Randament mediu - 42,3 q/ha , m randamentul maxim de 79,3 c/ha a fost obținut la Shchuchinsky GSU în 2001. Rezistent moderat la adăpostire și secetă. Rezistent la boli. Cerințe mari la condițiile de creștere. Reactivitate mare la fungicide. Sensibilitate medie la erbicide.
2.2 Metode de cercetare
Metode de cercetare - experiment, metoda comparatiei.
Experiența sa bazat pe următoarele opțiuni:
1) control (semințe fără tratament);
2) tratarea semințelor cu unde de 660 nm timp de 15 min;
3) tratarea semințelor cu unde de 660 nm timp de 30 min;
4) tratarea semințelor cu valuri de 775 nm timp de 15 minute
5) tratarea semințelor cu unde de 775 nm timp de 30 min.
În opțiunile 2-5, puterea expunerii laser (P) este de 100 mW.
Tratarea semințelor a fost efectuată pe sisteme laser (Figura 2.2).
Repetarea experienței de trei ori. Numărul de semințe în repetare - 20 buc.
În condiții de laborator, s-au determinat germinația și energia germinării semințelor. Pentru a face acest lucru, semințele culturilor de cereale au fost germinate la o temperatură de aproximativ 23 C timp de 7 zile.
Definiție înasemănări ale mugurilor de orz. Germinația a fost determinată pentru a stabili numărul de semințe capabile să producă răsaduri dezvoltate normal. La răsadurile dezvoltate în mod normal, rădăcina germinativă trebuie să aibă cel puțin jumătate din lungimea seminței. Pentru a calcula germinația semințelor dintr-o probă, se însumează numărul semințelor germinate normal când se ia în considerare germinația și numărul total al acestora se exprimă în %. În cursul acestui experiment, răsadurile au fost numărate cantitativ din aceleași locuri în a 7-a zi.
Determinarea energiei de germinare. Energia germinativă a fost determinată într-o analiză cu germinare, dar semințele germinate în mod normal au fost numărate în a 3-a zi.
La răsadurile dezvoltate în mod normal, rădăcina germinativă trebuie să aibă cel puțin lungimea sau diametrul seminței și, de obicei, cu peri de rădăcină, iar vlăstarul trebuie să aibă cel puțin jumătate din lungimea seminței. Acele specii care germinează cu mai multe rădăcini (orz, grâu, secară) trebuie să aibă cel puțin două rădăcini.
3. Efectul iradierii cu laser asupra vitezei de creștere a semințelor de orz
În urma studiului, s-a stabilit caracterul selectiv al efectului laser asupra ritmului de creștere a semințelor de orz, și anume energia de germinare și germinare. De regulă, starea semințelor determină cantitatea și calitatea recoltei.
Energia germinativă caracterizează prietenia și viteza germinării semințelor. Energia germinativă este procentul de semințe germinate în mod normal dintr-o probă prelevată pentru analiză.
Rezultatele cercetării noastre au arătat (Figura 3.1) că energia de germinare a semințelor de orz a fost cea mai mare atunci când au fost expuse la iradierea laser la o lungime de undă de 775 nm timp de 30 de minute. Comparativ cu controlul, a crescut cu 54% și a ajuns la 54%.
Semințele iradiate cu aceeași lungime de undă, doar pentru 15 minute, au avut o energie de germinare mai mică - 27%. Aceasta este mai mică decât rezultatele controlului de 1,3 ori.
Semințele iradiate cu o lungime de undă de 660 nm au avut o energie de germinare mai mică atunci când au fost iradiate timp de 30 de minute. Comparativ cu controlul, aceasta a scăzut cu 77% și sa ridicat la 8%. Când a fost iradiat cu aceeași lungime de undă, dar timp de 15 min, acest indicator a scăzut și el față de control cu 46% și a ajuns la 19%.
Germinarea semințelor este unul dintre indicatorii importanți ai calităților lor de semănat. O scădere a germinării chiar și cu 10-20% duce la o scădere de două trei ori a randamentului.
În timpul cercetării s-a stabilit efectul advers al tratamentului cu laser asupra germinării de laborator a semințelor de orz (Figura 3.2).
Cel mai deprimant a fost tratamentul cu unde cu lungimea de 660 nm timp de 30 min. În această variantă, comparativ cu martor (85%), rata de germinare a scăzut cu 75% și s-a ridicat la 21%. Când semințele sunt iradiate cu aceeași lungime de undă, dar timp de 15 minute, se observă o creștere a germinației, dar nu depășește valoarea de control. Acest indicator este mai mic decât controlul cu 18% și se ridica la 70%.
Tratamentul semințelor cu valuri de 775 nm a redus germinația acestora cu 33% (expunere 15 min) și 25% (expunere 30 min) comparativ cu martor.
Astfel, tratamentul cu laser nu a avut un efect pozitiv nici asupra energiei germinative a semințelor de orz cv. Tratamentul cu raze de 660 nm timp de 30 de minute a avut cel mai deprimant efect asupra germinării semințelor.
Concluzie
Astfel, după ce am studiat literatura pe această temă, putem trage următoarele concluzii:
1. Tratamentul înainte de însămânțare a semințelor cu substanțe chimice este asociat cu costuri ridicate cu forța de muncă și cu o capacitate scăzută de fabricație a procesului. În plus, utilizarea pesticidelor în scopul dezinfectării semințelor dăunează mult mediului.
2. Sub influența câmpului electromagnetic, forțele sunt mobilizate și rezervele de energie ale organismului sunt eliberate, procesele fiziologice și biochimice sunt activate în stadiile incipiente ale germinării semințelor, există o creștere a proceselor intra-metabolice și o creștere constantă. în energia germinativă, germinația, puterea, creșterea inițială, supraviețuirea primăvara-vară, care sunt favorabile afectează întreaga perioadă ulterioară de dezvoltare a plantei. Cu toate acestea, nu au primit o distribuție largă, deși sunt mai avansate din punct de vedere tehnologic, sigure pentru mediu și mult mai ieftine în comparație cu metodele chimice. Unul dintre motivele acestei situații este acela că metodele existente de tratare a semințelor cu radiații nu dau rezultate constant ridicate. Acest lucru se datorează modificărilor condițiilor externe, eterogenității materialului de semințe și cunoașterii insuficiente a esenței interacțiunii celulelor de semințe cu câmpurile electromagnetice și sarcinile electrice.
3. Metoda de tratament cu laser are o serie de avantaje față de metodele fizice și chimice de tratare a semințelor înainte de însămânțare:
Îmbunătățirea calității produselor agricole (creșterea conținutului de zaharuri, vitamine, proteine și gluten);
· posibilitatea reducerii ratei de însămânțare cu 10-30% prin creșterea germinării în câmp a semințelor și intensificarea proceselor de creștere;
Inofensivitatea procesării pentru semințe și personalul de service;
creșterea rezistenței plantelor la deteriorarea diverselor boli;
Durata scurtă a impactului
· o creștere a germinării semințelor unor plante cultivate, a germinării în câmp și a numărului de tulpini productive și, ca urmare, a productivității (până la 13%).
Dezavantajele acestei metode includ:
· dependența eficienței iradierii preînsămânțate de condițiile meteorologice din perioada de vegetație;
· un impact negativ asupra unui număr de caracteristici economice ale plantelor, o scădere a intensității regimului respirator al plantelor;
· o creștere a dozei de tratament poate provoca moartea;
foarte scumpe și, prin urmare, nu sunt utilizate pe scară largă în economie.
4. Pe baza rezultatelor cercetării noastre, putem trage următoarele concluzii:
Tratamentul cu laser nu a avut un efect pozitiv asupra energiei de germinare a semințelor soiului de orz Yakub, cu excepția variantei cu utilizarea razelor cu lungimea de undă de 775 nm timp de 30 de minute. În această variantă, s-a înregistrat o creștere a E ave cu 54% comparativ cu martor.
Utilizarea tratamentului cu laser cu o putere de 100 mW, indiferent de lungimea de undă și expunere, a redus germinarea semințelor de orz în condiții de laborator. Tratamentul cu raze de 660 nm timp de 30 de minute a avut cel mai deprimant efect asupra germinării semințelor.
Lista surselor utilizate
1. Atroşcenko, E.E. Efectul tratamentului semințelor cu unde de șoc asupra caracteristicilor morfofiziologice și a productivității plantelor: Ph.D. dis…. cand. bio. Științe: VAK 03.00.12. - M., 1997.
2. Veselova, T.V. Modificări ale stării semințelor în timpul depozitării, germinării și sub influența factorilor externi (radiații ionizante în doze mici și alte efecte slabe), determinate prin metoda luminiscenței întârziate: autor. dis…. dr. bio. Științe: 03.00.02-03. - M., 2008.
3. Danko, S.F. Intensificarea procesului de malțuire a orzului prin acțiunea sunetului de diferite frecvențe: dis…. cand. acestea. Științe: VAK RF. - M., 2001.
4. Eskov, E.K. Influența tratamentului semințelor de porumb cu pulbere ultrafină de fier asupra dezvoltării plantelor și a acumulării de elemente chimice în ele / E.K. Eskov // Agrochimie, Nr. 1, 2012. - P. 74-77.
5. Kazakova, A.S. Influența tratamentului înainte de însămânțare a semințelor de orz de primăvară de către un câmp electromagnetic de frecvență variabilă asupra calităților lor de semănat. / LA FEL DE. Kazakova, M.G. Fedorishchenko, P.A. Bondarenko // Tehnologie, agrochimie și protecția culturilor agricole. Culegere interuniversitară de lucrări științifice. Zernograd, 2005. Ed. RIO FGOU VPO ACHGAA. - S. 207-210.
6. Ksenz, N.V. Analiza efectelor electrice și magnetice asupra semințelor / N.V. Ksenz, S.V. Kacheishvili // Mecanizarea și electrificarea agriculturii. - 2000. - Nr. 5. - S. 10-12.
7. Melnikova, A.M. Efectul iradierii cu laser asupra germinării semințelor și dezvoltării răsadurilor / Melnikova A.M., Pastukhova N. // Ecologie. Siguranța la radiații. Probleme socio-ecologice. - Universitatea Tehnică de Stat Donbass.
8. Neshchadim, N.N. Studiu teoretic al efectului tratamentului semințelor și culturilor cu substanțe de creștere, câmp magnetic, iradiere cu laser asupra producției și calității produsului, recomandări practice; experimente cu grâu, orz, alune și trandafiri: autor. dis…. dr. Științe agricole: Universitatea Agronomică Kuban. - Krasnodar, 1997.
9. Novitskaya, G.V. Modificări ale compoziției și conținutului de lipide din frunzele tipurilor de ridichi orientate magnetic sub influența unui câmp magnetic constant slab / G.V. Novitskaya, T.V. Feofilaktova, T.K. Kocheshkova, I.U. Yusupova, Yu.I. Novitsky // Fiziologia plantelor, V. 55, nr. 4. - S. 541-551.
10. Novitskaya, G.V. Influența unui câmp magnetic alternant asupra compoziției și conținutului de lipide la răsadurile de ridichi / G.V. Novitskaya, O.A. Tserenova, T.K. Kocheshkova, Yu.I. Novitsky // Fiziologia plantelor, V. 53, nr. 1. - S. 83-93.
11. Novitskaya, G.V. Influența unui câmp magnetic constant slab asupra compoziției și conținutului de lipide al frunzelor de ceapă de diferite vârste / G.V. Novitskaya, T.K. Kocheshkova, Yu.I. Novitsky // Fiziologia plantelor, V. 53, nr. 3. -
p. 721-731.
12. Tratarea semințelor - protecție împotriva bolilor și garantarea recoltei // ChPUP „Biohim” URL: http://biohim-bel.com/obrabotka-semyan (Accesat: 20.03.2013).
13. Rakhmankulova, Z.F. Influența tratamentului de preînsămânțare a semințelor de grâu cu acid salicilic asupra conținutului său endogen, activității căilor respiratorii și echilibrului antioxidant al plantelor / Z.F. Rakhmankulova, V.V. Fedyaev, S.R. Rakhmatullina, S.P. Ivanov, I.G. Gilvanova, I.Yu. Usmanov // Fiziologia plantelor, vol. 57, nr. 6, p. 835-840.
Documente similare
Sistemul de producție de semințe de ierburi perene în Republica Belarus. Caracteristicile morfologice și biologice-ecologice ale pajiștii de iarbă albastră. Influența tratamentului semințelor cu regulatori de creștere asupra germinării câmpului și supraviețuirii semințelor, asupra productivității semințelor.
teză, adăugată 07.10.2013
Dormința semințelor și condițiile pentru depășirea acesteia. Condițiile fizico-geografice, solului și climatice ale regiunii Irkutsk. Caracteristicile ecologice și morfologice ale plantelor studiate. Eficiența economică a utilizării albitului pentru îmbunătățirea germinării semințelor.
teză, adăugată 14.10.2011
Caracteristici ale creșterii și dezvoltării boabelor de soia. Boli și dăunători. Regulatori ai creșterii și dezvoltării plantelor, ca element de tehnologie care crește rezistența plantelor la stres. Caracteristici de creștere și dezvoltare a soiului de soia Vilana. Tratamentul de pre-semănat al semințelor cu regulatori.
teză, adăugată 26.02.2009
Descrierea necesității de zinc pentru creșterea normală a unui număr mare de specii de plante superioare. Studiul influenței Znului asupra gradului de germinare a semințelor de floarea soarelui. Măsurarea conținutului de clorofilă. Determinarea capacității de absorbție a sistemului radicular.
raport de practică, adăugat la 27.08.2015
Randamentul de soia în regiunea Kaluga. Eficiența simbiozei leguminoase-rhizobium. Conținutul de proteine din boabele de soia. Randamentul semințelor de soia în funcție de tipul de preparat și metoda de tratament cu regulatori de creștere. Înmuierea semințelor în soluție de fusicoccină.
articol, adăugat 08.02.2013
Ciupercile din genul Fusarium ca agenți patogeni ai peste 200 de specii de plante cultivate. Surse de infecție primară: semințe, sol, resturi vegetale. Caracteristicile metodei de germinare a semințelor. Importanța ciupercilor micorizice în alimentația plantelor superioare.
teză, adăugată 04.11.2012
Cercetarea valorii economice și a caracteristicilor biologice ale orzului de primăvară. Rolul nutriției minerale pentru orz. Analiza efectului îngrășămintelor și produselor de protecție a plantelor asupra randamentului, compoziției chimice și calității culturii, asupra dezvoltării bolilor orzului.
lucrare de termen, adăugată 15.12.2013
Caracteristicile generale ale RRR. Efectul fitohormonilor asupra creșterii țesuturilor și organelor, formării semințelor și fructelor. Mecanismul de acțiune al fitohormonilor asupra stării de stres a plantelor, creșterea și morfogeneza acestora. Utilizarea fitohormonilor și a substanțelor active fiziologic.
lucrare de control, adaugat 11.11.2010
Caracteristicile cultivării orzului de primăvară, caracteristicile sale biologice, în special cultivarea solului și a semințelor. Ratele de consum de pesticide pentru tratarea culturilor de orz de la dăunători. Esența și scopul cerințelor grele, agrotehnice.
lucrare de termen, adăugată 01/04/2011
Procesul de prelucrare post-recoltare a cerealelor. Ventilația activă a cerealelor și semințelor. Principalele tipuri de hambare din întreprinderile agricole. Performanța operațională a mașinii de curățare secundară MVU-1500. Tehnologia prelucrării în orz perlat.
slide 1
Efectul tratamentului semințelor de fasole cu soluții chimice asupra creșterii și dezvoltării plantelorslide 2
 Scopul studiului: acela de a afla efectul stimulant al tratamentului semințelor cu diverse substanțe chimice asupra dezvoltării plantelor de fasole. Ipoteza: Tratamentul semintelor are un efect stimulativ asupra dezvoltarii plantelor
Scopul studiului: acela de a afla efectul stimulant al tratamentului semințelor cu diverse substanțe chimice asupra dezvoltării plantelor de fasole. Ipoteza: Tratamentul semintelor are un efect stimulativ asupra dezvoltarii plantelor
slide 3
 Obiectivele cercetării: să dea o descriere științifică a efectului stimulator al diferitelor substanțe chimice asupra dezvoltării plantelor; stăpânește metodologia experimentului pentru a identifica efectul stimulator al diferitelor substanțe chimice asupra dezvoltării plantelor; utilizarea metodei propuse pentru a investiga efectul stimulator al a șase substanțe chimice asupra dezvoltării plantelor de fasole; trageți concluzii pe baza rezultatelor obținute cu privire la efectul stimulator al substanțelor chimice asupra creșterii și dezvoltării plantelor.
Obiectivele cercetării: să dea o descriere științifică a efectului stimulator al diferitelor substanțe chimice asupra dezvoltării plantelor; stăpânește metodologia experimentului pentru a identifica efectul stimulator al diferitelor substanțe chimice asupra dezvoltării plantelor; utilizarea metodei propuse pentru a investiga efectul stimulator al a șase substanțe chimice asupra dezvoltării plantelor de fasole; trageți concluzii pe baza rezultatelor obținute cu privire la efectul stimulator al substanțelor chimice asupra creșterii și dezvoltării plantelor.
slide 4
 Relevanța studiului: producția modernă a culturilor nu poate face fără tehnici speciale care măresc randamentul plantelor, îmbunătățesc creșterea și dezvoltarea acestora și protejează împotriva bolilor și dăunătorilor. În prezent, tratamentul semințelor înainte de însămânțare este utilizat în practică. Cu toate acestea, nu există informații complete despre care substanțe chimice și cum afectează acestea semințele anumitor plante, cum afectează tratamentul semințelor diferitele faze ale plantei. În acest sens, tema studiului nostru este relevantă.
Relevanța studiului: producția modernă a culturilor nu poate face fără tehnici speciale care măresc randamentul plantelor, îmbunătățesc creșterea și dezvoltarea acestora și protejează împotriva bolilor și dăunătorilor. În prezent, tratamentul semințelor înainte de însămânțare este utilizat în practică. Cu toate acestea, nu există informații complete despre care substanțe chimice și cum afectează acestea semințele anumitor plante, cum afectează tratamentul semințelor diferitele faze ale plantei. În acest sens, tema studiului nostru este relevantă.
slide 5
 STUDIU PRACTIC Prima probă - 1% soluție de sare alimentară A 2-a probă -% soluție de glucoză A 3-a probă -1% soluție de sifon de băut A 4-a probă -1% soluție de acid boric A 5-a probă -1% soluție de permanganat de potasiu A 6-a probă - apă.
STUDIU PRACTIC Prima probă - 1% soluție de sare alimentară A 2-a probă -% soluție de glucoză A 3-a probă -1% soluție de sifon de băut A 4-a probă -1% soluție de acid boric A 5-a probă -1% soluție de permanganat de potasiu A 6-a probă - apă.
slide 6

Slide 7

Slide 8

Slide 9
 Rezultatele studiului au arătat că multe substanțe chimice sunt regulatoare de creștere a plantelor; unele substanțe chimice au un efect inhibitor. În acest caz, este o soluție de acid boric; efectul stimulator al substanțelor chimice se manifestă în diferite stadii de dezvoltare a plantelor de fasole; sarea comestibilă afectează rata de coacere a fructelor; consumul de sifon afectează rata de apariție a rădăcinii germinale, dinamica răsadurilor; permanganatul de potasiu afectează înălțimea plantelor; glucoza afectează randamentul.
Rezultatele studiului au arătat că multe substanțe chimice sunt regulatoare de creștere a plantelor; unele substanțe chimice au un efect inhibitor. În acest caz, este o soluție de acid boric; efectul stimulator al substanțelor chimice se manifestă în diferite stadii de dezvoltare a plantelor de fasole; sarea comestibilă afectează rata de coacere a fructelor; consumul de sifon afectează rata de apariție a rădăcinii germinale, dinamica răsadurilor; permanganatul de potasiu afectează înălțimea plantelor; glucoza afectează randamentul.